Lecture
The nature of rhythms is traditionally correlated with the oscillatory properties of neurons. So, the behavior of a single neuron is described by the Hodgkin-Huxley equations (Hodgkin, 1952):
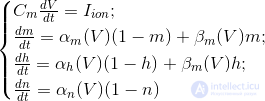
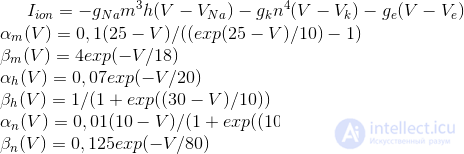
These equations determine the autowave process, that is, they prescribe a neuron to generate pulses. The Hodgkin-Huxley model is difficult to model. Therefore, there are many of its simplifications that preserve the basic generating properties. The most popular models are Fitzhugh-Nagumo (Fitzhugh, 1961), Morris-Lecar (Morris C., Lecar H., 1981), Hindmarsh-Rose (Hindmarsh JL, and Rose RM, 1984). Many models, for example, Hindmarsh-Rose, allow you to simulate both bundling activity and isolated commissures (figure below).
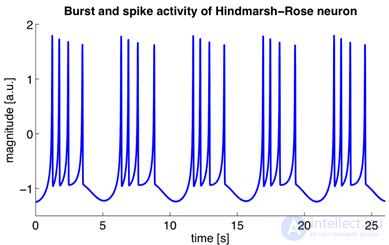
Bundle activity and adhesions arising from the Hindmarsh-Rose neuron simulation
By combining neurons that generate their own impulses, in constructions resembling the structure of a real cortex, it is possible to reproduce various effects characteristic of the group activity of real neurons. For example, you can achieve global synchronization of neural activity or cause the appearance of waves. The most famous models are Wilson-Cowan (HR Wilson and JD Cowan, 1972) and Kuromoto (Kuramoto, 1984).
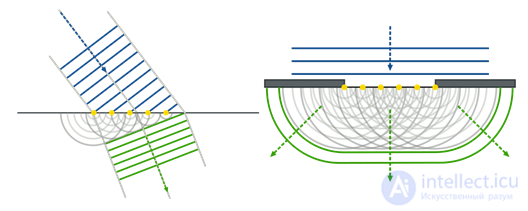
An electroencephalogram captures echoes of joint activity of neurons, but it is clear that this activity has a certain space-time organization. The methods of optical observation of the activity of the cortex make it possible to see it live. An experimental animal is exposed to a portion of the cortex and a special dye sensitive to changes in electrical potential is introduced. Under the influence of total oscillations of the membrane potential of neurons, such a dye changes its spectral properties. And although these changes are extremely small, they can nevertheless be recorded, for example, with the help of a diode array that performs the role of a high-speed video camera. Optical methods do not allow you to look deep into the cortex and trace the activity of individual neurons, but they provide an opportunity to get a general idea of the course of wave processes on its surface.
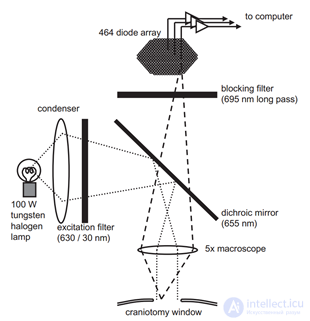
Optical fixation of bark activity (Michael T. Lippert, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu, 2007)
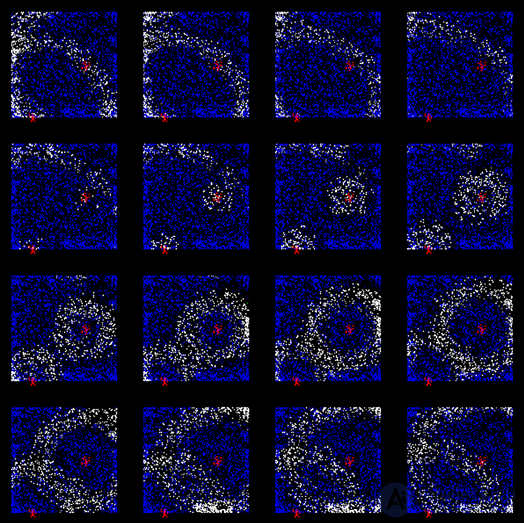
It turned out that the rhythms of the brain correspond to the waves that occur in point sources and propagate through the cortex like circles in water. True, the wave front on the water diverges strictly in a circle, and the wave front of the activity of brain neurons can spread in a more complicated way. The figure below shows the patterns of wave propagation in the 5 millimeter section of the rat cortex.
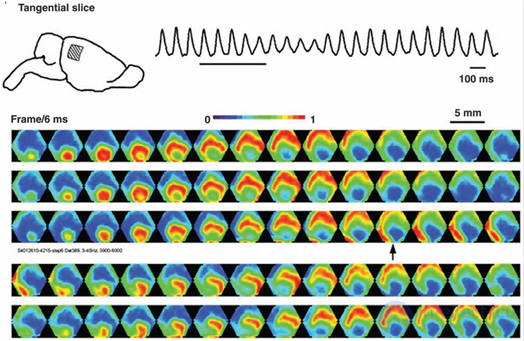
Picture of wave propagation of activity in the rat cortex. The potential is shown by a gradient from blue to red. 14 frames with an interval of 6 milliseconds cover one wave propagation cycle (84 milliseconds - 12 Hz) (Michael T. Lippert, Kentaroh Takagaki, Weifeng Xu, Xiaoying Huang, Jian-Young Wu, 2007)
Very interesting and illustrative wave activity videos are given in (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y.Wu, 2007). There it is shown that the waves can condense, reaching the boundary of the crustal zone, can be reflected from another zone and create an oncoming wave, they can spread with double spirals and create vortices.
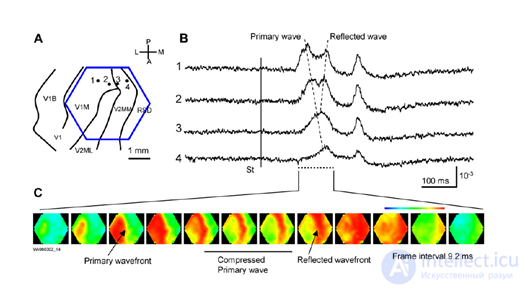
Compression and wave reflection in the primary visual cortex (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y. Wu, 2007)
Below are videos from the same work (W.-F. Xu, X.-Y. Huang, K. Takagaki, and J.-Y.Wu, 2007):
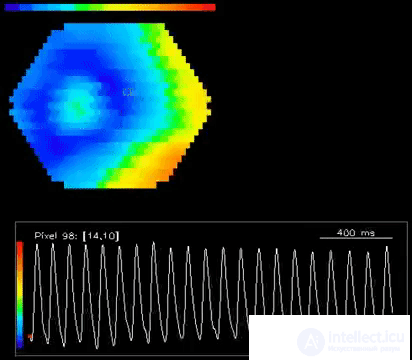
Using the models of oscillating neurons, it is possible in computer experiments to obtain similar wave patterns. But by itself, such a pulsation does not make much sense. It is quite reasonable to assume that activity waves are a mechanism for transferring and processing information. But attempts in traditional models to explain the nature of this mechanism did not yield tangible results. It seems logical to assume that the information is encoded by the frequency and phase characteristics of neural signals. But this leads to the need to take into account interference processes and raises more questions than clarifies.
There is a hypothesis that the waves “scan” the cortex, reading information from it for its subsequent transmission (Pitts W., McCulloch WS, 1947). It looks quite sensible, at least with respect to alpha waves and the primary visual cortex. The authors of this hypothesis, McCulloch and Pitts, emphasized that scanning allows us to explain one important phenomenon. The fact is that the information from the primary visual cortex is projected further along the fibers of the white matter through axon bundles, which are clearly insufficient in their volume to simultaneously transmit the entire state of this zone of the cortex. So, they concluded, scanning allows you to use not only the spatial but also the time code, which ensures the sequential transmission of the required information.
This problem of narrow projection beams exists not only for the primary visual cortex, but also for all other areas of the brain. The number of fibers in the projection paths is much less than the number of neurons that form the spatial pattern of activity. The links between the zones are clearly not able to simultaneously transmit the entire spatially distributed signal, which means that an understanding of how compression and decompression of the information transmitted through them is required. The assumption of scanning, although it does not answer the question about the coding mechanism itself, nevertheless allows us to ask ourselves the right questions.
Our model in explaining brain rhythms is based on the above description of the nature of metabotropic activity. Using the idea that not only neuronal synapses are involved in the process of creating rhythms, but also metabotropic receptive clusters, allows us to obtain a result that is qualitatively different from all classical theories. But before proceeding to the description of this, I want to make a warning. I will deliberately describe simplified idealized models, matching them with certain processes inherent in the real brain, but not claiming that the brain works that way. Our task is to show the basic principles, understanding that evolution has gone far ahead and their true implementation is much more cunning. You can draw an analogy with the development of computing. A modern computer is quite complicated, and if we begin to describe the basic principles of classical computing devices, it turns out that in their pure form it is almost impossible to meet them in modern systems. Basic view - the processor reads the program and data from memory, performs the actions prescribed by the program on the data, and writes the results back to memory. And now add to this the use of different levels of cache, multithreading, hyperthreading, parallel computing using local, group and shared memory, and the like. And it turns out that it is difficult to find in the real computer literal observance of simple rules. Actually, all this must be taken into account when comparing the following description with the work of the real brain.
So, let's take a model of the area of the cortex, on which we will create a compact pattern of induced activity. For now let's leave the question how this pattern came about. We simply assume that there are elements on which there is a constant pulse signal. In the image below, the neurons forming such a pattern are marked in red. In real bark, this corresponds to axons transmitting packet activity, that is, issuing a series of spikes with a high frequency. These axons may belong to neurons of the same zone of the cortex that are in a state of induced excitation, or they may be projection fibers coming from other parts of the brain.

Invoked Activity Pattern
Now let the free neurons generate rare random pulses. At the same time, we impose a condition that for a random spike a certain level of environmental activity is required. This means that random spikes can occur only in the vicinity of already active neurons. In our case, they will appear in the neighborhood of the pattern of evoked activity. In the image below, random spikes are shown in green.
Usually, all neuron activity that is not induced is called spontaneous or background. This is not very successful, as it adjusts to the perception of all this activity as random. Further we will show that, in general, the background activity is strictly predetermined and not at all accidental. And only a small part of it is really completely random spikes. We have now created such random spikes around an active pattern.
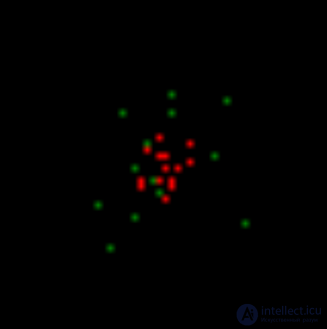
The first step is modeling. Spontaneous activity on the background of induced activity
Let us model metabotropic receptive clusters on our formal neurons. To do this, we will allow neurons to memorize, when necessary, a picture of the activity of the nearest environment. And we will not limit ourselves to one picture for one neuron, as it would be, we memorize at synapses, but let each neuron store a multitude of such pictures.
Let us make it so that the presence of our own random spike and high environmental activity will every time be a reason to memorize the picture of surrounding impulses (figure below). Further, our neurons will give single spikes every time one of these memorized local pictures is repeated. In order not to confuse them with random spikes, we will call these spikes wave.

The area of activity fixation for one of the neurons that issued a spontaneous spike (it is in the center of the square). When modeling, for simplicity, square receptive fields were used, which, although not consistent with the tracking fields of real neurons, does not qualitatively affect the result.
As a result, in the next modeling step (figure below) we will get a picture in which there will be the same as before, neurons with induced activity (red), neurons that reacted to the local images (white) memorized in the previous step, and neurons that generated spontaneous spike (green).
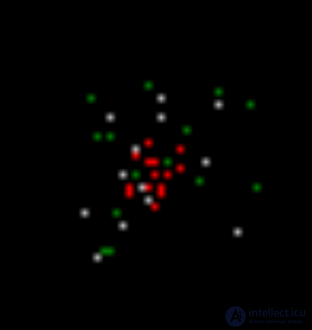
The second step is modeling. Red - induced activity, white - wave activity, green - spontaneous activity
Repeating the steps of the simulation, we get the activity spreading through the cortex with a certain unique randomly created pattern.
Now we introduce the neuron fatigue condition. Let us make it so that after several metabotropic (wave) spikes, neurons will lose the ability to generate new impulses for the time needed for relaxation. This will lead to the fact that the activity will spread not a continuous area, but pulsating waves, scattering from its source.
To prevent “clogging,” we block spontaneous activity in areas where there are a large number of relaxing neurons. As a result, we get a ring of neurons with wave activity, running away from the pattern of evoked activity (figure below).

Wavefront propagation. Blue - neurons in a state of relaxation
The resulting wave as it moves away from its center will lose the density of the front and sooner or later it will attenuate. In the figure below you can trace the first cycle of her life.
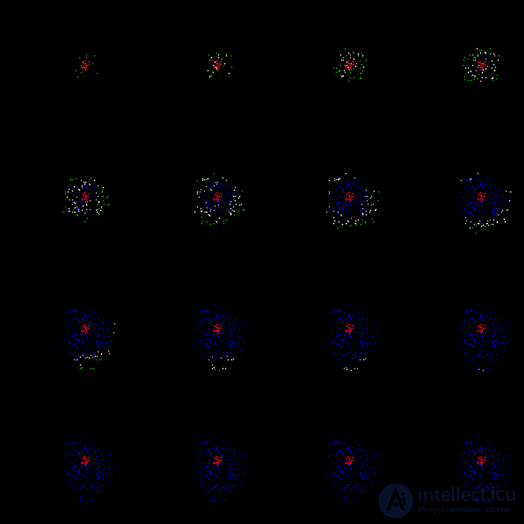
The first cycle of propagation of the wave id
After the relaxation period passes, a new wave will start. Now it will be picked up by neurons that have already been trained, and will spread a little further. With each new cycle, the wave will expand its propagation limits until it reaches the edges of the zone. To prevent an unlimited increase in the density of the wave front, we introduce one more restriction: we prohibit spontaneous activity for neurons when the total activity around them exceeds a certain threshold.
After some time, we will get a cortex trained to propagate a unique in its pattern wave of activity corresponding to the initial pattern of evoked activity originally given (figure below).
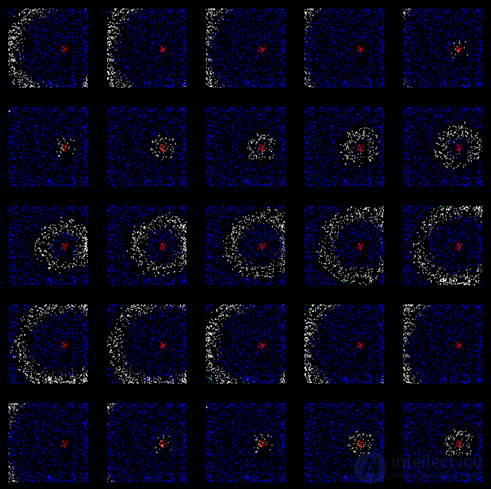
Wave propagation along the already trained cortex (after 200 training cycles)
Note: when the cortex is already trained to spread a wave from a particular pattern, spontaneous (random) spikes disappear. Such an "experienced" wave is formed by wave, non-random spikes. Random spikes appear only when training, when the wave front reaches still untrained or poorly trained areas. At this point, spontaneous pulses create a random continuation of the wave pattern, which is immediately remembered on the surface of the neurons involved in this wave. But as soon as the training ends, the wave from the pattern familiar to the core begins to spread along the already created wave pattern, each beat repeating exactly the same non-random pattern in its path.
Learning the cortex does not have to be phased. When simulating the selection of parameters, it is possible to ensure that the wave that originated is self-sufficient. That is, so that it does not fade at the first bars, but immediately spread to the whole space (figure below).
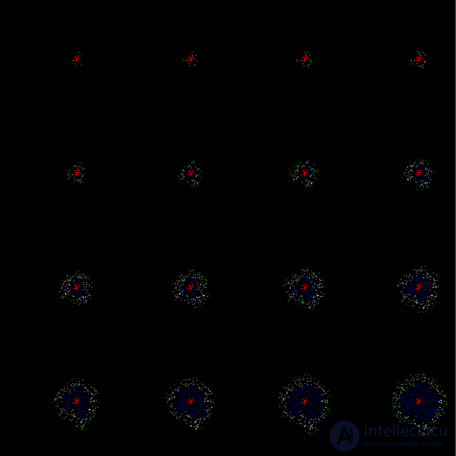
An example of a continuous on the first cycle of the propagation of a training wave
Since the width of the wave front is constant, its area increases with distance from the source. This corresponds to the fact that more and more neurons participate in the dissemination of activity. If we trace the total potential created by them, then we get graphs that resemble what we see on the entsifologramma (figure below).
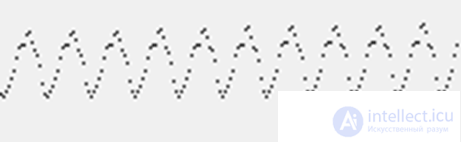
Graph of rhythmic activity of a single source model
Please note that the appearance of rhythms observed on such “encephalograms” is not the rhythm of the “respiration” of the cortex. That is, it is not the synchronization of joint bursts of activity, as is customary in most existing models, but a change in the number of neurons involved in the propagation of diverging waves. If new waves are radiated even before the fronts of previous waves disappear, their addition will give a smoother picture. With a certain selection of parameters on the model, situations are reproduced where wave propagation is practically not accompanied by total rhythmic activity. This can be correlated with the fact that about 10 percent of people on the electroencephalogram wave activity can not be traced.
If you now take a new pattern of evoked activity, then the crust will create waves that extend from it. Moreover, such a cortex is trained to create waves for any stable pattern of evoked activity. Due to the fact that the same neuron can store many local images, it can be part of many different waves at once, corresponding to different patterns. If we want a greater uniqueness of the waves, then as the neuron accumulates the stored local images, it reduces the likelihood of its spontaneous spike.
For stored local images, it makes sense to introduce a consolidation mechanism. That is, do not fix the image immediately for ages, but set the time during which the image should repeat a certain number of times. Since wave propagation training and the formation of sustainable patterns of evoked activity are parallel processes, consolidation can erase the traces of unsuccessful learning.
But the most important thing in all this is the uniqueness of each of the received wave histories. In any place through which the wave passes, she creates a single pattern, characteristic only for her. This means that if there is a pattern of evoked activity on the surface of the cortex that encodes an event, then the wave caused by it will spread information about it throughout the entire space of the cortex. In each particular place, this wave will create its own unique pattern, unique only for this event. Any other event will create in this place a different pattern associated with its wave.This means that, being in any place, we can tune in to the “reception” of the wave from any particular event and fix when it happens after the arrival of the wave with the corresponding pattern.
In our model, it turns out that the fundamental properties of the cortex are:

Comments
To leave a comment
Logic of thinking
Terms: Logic of thinking