This series of articles describes a wave model of the brain that is seriously different from traditional models. I strongly recommend that those who have just joined begin reading from the first part.
Let's continue the conversation about neuron detectors. Suppose that some information is projected onto the zone of the cortex through wave tunnels. Each of the projection beams are axons of neurons located on the zone that sends this information. The projection is removed from a small area of bark. The fibers of the projection beam, in fact, transmit the wave patterns passing through this section. The place of the host cortex, where the projection falls, becomes the source of the waves. These waves carry the same information on the receiving zone of the cortex as the waves on the source zone.
If we adjust the weights of a neuron to recognize a specific wave pattern passing through its receptive field, then we will turn it into a detector that is triggered at the moment when the characteristic combination of identifiers for it appears.
If we teach several neurons located in relative proximity to each other to detect the same wave pattern, then we will receive not a single neuron detector, but a detector pattern. The response of a single neuron detector to a characteristic stimulus is a packet of impulses of induced activity. The response of a detector pattern is the induced activity of a group of neurons that form a certain pattern. Naturally, such a pattern of evoked activity will begin to train the cortex to distribute its unique identifier.
It turns out that when a bark zone receives a description consisting of certain signs, its detector patterns react to familiar combinations and generate their own waves, which carry information about new signs recognized by this zone. Thus, the zone of the cortex first detects statistical patterns in the attribute pattern arriving at it and generates patterns of detectors corresponding to these patterns, and then, receiving descriptions in only signs, it generates its own description in those signs that were previously detected by this zone.
The logic of such a transformation of neural descriptions can be compared with the logic of verbal descriptions, which is not surprising, since these phenomena are closely related. Suppose we see different beasts. Each of them we can describe a set of signs. For example, we see a beast with a description: big, brown, shaggy. Not knowing what kind of animal it is, we can call it that: big-brown-shaggy. The combination of features forms the identifier of this animal. If this beast occurs to us often, then we can fix the pattern that these signs are found together, and give this combination its own name, for example - the bear. Further, we can operate on the concept of "bear" as independent. At the same time, we have the opportunity, if necessary, to restore all the original features of the concept of a "bear".
Speaking of the real world, it is convenient to use the term "phenomenon". Its convenience is in its community. This term describes not only objects, but also the processes occurring with them. Any picture we perceive can be called a phenomenon. Every phenomenon we encounter gives rise to a description. Description is a combination of the pattern of activity of neuron detectors that form patterns and the identification waves generated by them.
It is possible to call the pattern of neuron-detectors operating on one image, and the corresponding identification wave, a concept. Then we can say that any phenomenon is represented by the brain as a set of descriptions consisting of concepts. For each zone of the cortex description is based on its own set of concepts.
When the neurons-detectors isolate any new pattern or fix a new combination of features so that this creates a pattern of such detectors that are capable of generating their own unique wave, we can talk about the formation of a new concept. The interpretation of this concept can be those real-world events that have created such a combination of signs, and to which the corresponding neurons are now responding.
Pattern formation. Reverse projection
The pattern of neuron detectors tuned to a common stimulus allows you to launch your own unique identifier wave. This corresponds to the recognition of the image and the inclusion of the advising concept in the description of what is happening. For an identifier to be unique, it is sufficient that the neuron detectors create their own pattern, unlike any of the wave patterns passing through their location. We describe a simple model that creates such a pattern.
Let us return briefly to the description of the mechanisms of neural interaction. The main way of transmitting signals from one neuron to another is the release of neurotransmitters from the axon terminal of the transmitting neuron into the synaptic cleft. On the surface of the neuron that receives the signal, are receptors that respond to a specific neurotransmitter, which is a ligand for it. Some receptors are located in the synapses and are responsible for synaptic transmission, some are outside the synapses and are responsible for wave activity.
There are a lot of neurotransmitters, including neuroactive peptides, there are more than fifty of them. The most common activating neurotransmitter is glutamate, the suppressor is gamma-aminobutyric acid (GABA). The ability to activate or suppress is not a property of the neurotransmitter itself, but is determined by the type of receptor interacting with it. In order for the signal transmission to take place, it is necessary that the neurotransmitter ejected by the axon and the receptors that receive it match each other.
Until the end of the 80s of the last century, it was believed that each neuron ejected the same mediator in all terminals of its axon - the “Dale principle”. But later it was shown that there are neurons using various combinations of neurotransmitters.
Take for simulation neurons of two hypothetical types, working with one mediator, let's call it (A). The difference between the types of neurons will be in where they will have receptors sensitive to a common mediator (the table below).
| Synapses | Outside synapses | Axon |
| Type 1 | - | A | BUT |
| Type 2 | A | - | BUT |
Neurotransmitter characteristics of neurons. Receptor sensitivity and axon release The neurons of the first type do not have receptors sensitive to the mediator used in their synapses. But they have such receptors on the extra-synaptic part of the membrane. This means that these neurons are not capable of synaptic training on the patterns of activity that are created by neurons of both the first and second types, but they are able to learn how to propagate wave activity and be transmitters of the identification wave.
Let us now take the zone of the cortex, consisting of neurons of both types. For example, just mix them randomly. The presence of inclusions of neurons of the second type will not affect in any way the ability of the cortex to be trained to conduct waves of identifiers using neurons of the first type.
Suppose that we somehow determined the place on the crust where we want to create a detector pattern.
 Place chosen for training
Place chosen for training Suppose that we have solved all the issues related to training, and our task now is simply to fix the prototype. Let's do it in a way we already know. Let's force the second type of neurons, located in this area and unoccupied in training, to participate in the generation of random spikes. We set the spike probability to get as many active neurons as there are neurons-detectors that we need in the final pattern. Random spikes will create a unique pattern of neurons of the second type, localized in the selected area.
Next, we will do in accordance with the principle of Hebb, use the activity of neurons of the second type as a signal for their learning. We change the weights of their synapses in accordance with the wave pattern observed by them in their receptive fields. As a result, we get a pattern of neuron detectors tuned to the just passed set of identifiers. The repetition of the wave pattern will lead to the induced activation of all neurons in the pattern. The neurons of the first type will pick up this pattern and turn it into a propagating wave of a new identifier.
We got a very simple, but remarkable result. A certain combination of identifiers can generate its own unique identifier.
Now we will somewhat complicate the model. Add another mediator (B) and change the characteristics of the neurons (table below).
| Synapses | Outside synapses | Axon |
| Type 1 | B | A | BUT |
| Type 2 | A | B | B |
Characteristics of neurons using two mediators. Receptor sensitivity and axon release The behavior of this design will be much more interesting. Having created a detector pattern, neurons of the second type cannot propagate a wave from it through neurons of the first type, since their type of axon mediator does not coincide with the extra-synaptic sensitivity of neurons of the first type. But then these neurons will be able to spread the wave of their identifier themselves. In this case, it is interesting that the input and output activity of the cortex will be divided. If we somewhere in the side create an output wave tunnel from axons of neurons of the second type, then it will not transmit the entire wave pattern consisting, among other things, of a multitude of input features, but will transmit further only the description consisting of recognized concepts.
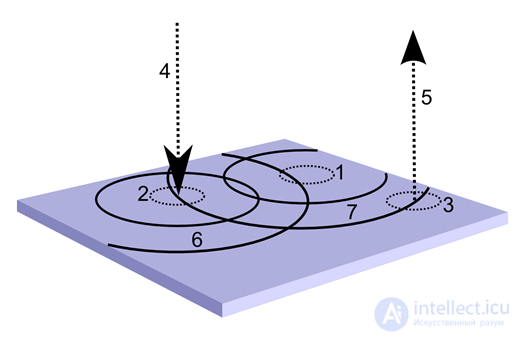 Changing the description on the bark zone. 1 is the selected training area, 2 is the region emitting the input identifier, 3 is the output area, 4 is the input wave tunnel, 5 is the output wave tunnel, 6 is the input wave of the identifier, 7 is the output wave of the identifier
Changing the description on the bark zone. 1 is the selected training area, 2 is the region emitting the input identifier, 3 is the output area, 4 is the input wave tunnel, 5 is the output wave tunnel, 6 is the input wave of the identifier, 7 is the output wave of the identifier But that is not all. In the same chosen area, we can conduct synaptic training of neurons of the first type. For them, the signal to learn will be their own wave activity. Since we have set the sensitivity of synapses of neurons of the first type only to the mediator (B), the pictures that they perceive will be the pattern of the same pattern that we previously trained on neurons of the second type. At the same time, the neurons-detectors of the first type themselves will line up in a pattern that repeats the wave of identifiers that took place at the time of training. This means that if we return the identifier of a concept that was previously detected by neurons of the second type to the cortex through the counter-wave tunnel, then spreading to the selected location, it will trigger the activity of the pattern of neuron detectors of the first type. And since these neurons repeat the pattern of a fragment of the source identifiers, their evoked activity will cause the corresponding identification waves to propagate through neurons of the first type.
Thus, we reproduced the rear projection mechanism. The bark in our model is able to broadcast information in both directions. In the forward direction, the integration of features and the description change to a more general one. In the opposite direction, having received the corresponding identifier, the cortex restores all the corresponding signs and broadcasts them down, if, of course, there is a corresponding projection connection.
We got a fairly simple construction: in one direction - we learned the concept, in the opposite direction - we restored the signs. But this simplicity of two-way communication is a unique feature for neural networks. Traditional networks are ideologically unidirectional, they cannot trigger a “back axon” signal, and this flaw cannot be cured by any tricks. At the same time, it is obvious that the real brain is capable of not only summarizing information, raising the description from level to level, making it more abstract, but also translating the description back, turning it into images we represent, internal speech, physical actions. Therefore, it is particularly gratifying that our design makes it possible to reproduce the reverse projection rather naturally, while remaining within the framework of the mechanisms accessible to the real brain.
If we force axons of neurons of one of the types to generate two mediators at once, then we will obtain the distribution of the corresponding identifiers in two layers at once. You can imagine the situation where it will be useful. In general, combinations of mediators and receptors can program the most unexpected behavior of the cortex. At the same time, it is pleasant that our core does not require any special topological configuration. Neurons form synaptic contacts with their surroundings, without worrying about adhering to any connection schemes. It seems that similar architectural principles are peculiar to real bark.

Comments