This series of articles describes a wave model of the brain that is seriously different from traditional models. I strongly recommend that those who have just joined begin reading from the first part.
The complete removal of the hippocampus makes it impossible to form new memories, which convincingly demonstrated the case of the HM patient. Disruptions in the work of the hippocampus can lead to Korsakov's syndrome, which also boils down to the inability to fix current events while maintaining the old memory. All this proves that the hippocampus plays a key role in the memory mechanism.
Traditional theories about the role of the hippocampus are built on the analogy of the brain and the computer. In such reasonings, the hippocampus plays the role of "operative memory", that is, the place where new memories accumulate. Then, presumably in a dream, these memories are transferred to the areas of the brain responsible for storing long-term memory. And although the mechanisms of such a transfer are incomprehensible, at least this helps explain why the hippocampal disruption blocks the formation of event memory.
Our model is fundamentally different from traditional models. We have no memory buffering in the hippocampus and its subsequent copying. The elements that make up the memories are immediately formed where it is appropriate for them to be. At the same time, the hippocampus simply creates a single identifier that unites the memory elements distributed over the space of the cortex. The wave model of the brain explains how this identifier spreads throughout the cortex. The presence of such an identifier makes it possible, by choosing the elements it has combined, to reproduce not an abstract picture, but a concrete image of the memorized event.
In 2010, interesting experiments on prosthetics of the rat hippocampus were made (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011). Experimental animals on both sides of the brain microelectrode arrays, exciting regions of CA3 and CA1, were introduced into the hippocampus. After the rats recovered after the operation, a series of tests began.
Rats were placed in a chamber with two retractable levers. The test consisted of three phases. At first, one of the levers was put forward at random. When the rat pressed this lever, a picture of the activity of the preselected hippocampal neurons was fixed (sample stage). Then the lever was removed, and there was a pause from 1 to 30 seconds in some series and up to 60 in others. After that both levers were put forward. When you press the lever opposite to the original sample, reinforcement was issued in the form of a drop of water. In case of an error, that is, pressing the same lever that was at the sample stage, the punishment followed - turning off the light for 5 seconds. As a result, a graph of the success of attempts against the delay time was compiled (figure below).
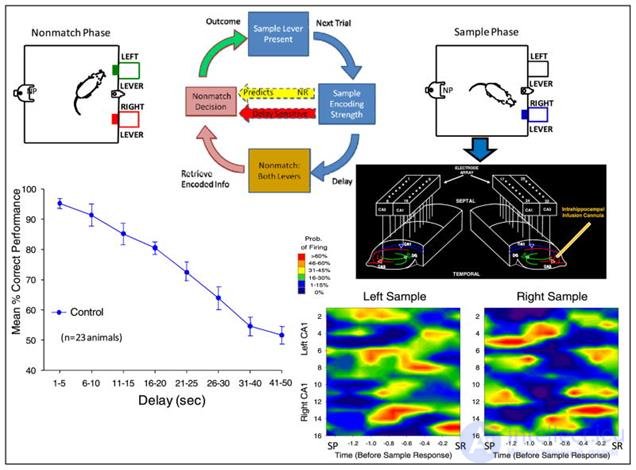 Control measurements with natural hippocampus activity (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Control measurements with natural hippocampus activity (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011) Based on the recorded signals, a predictive model was made that predicted which signal should be waited on CA1 depending on the signal on CA3. In a series of experiments, forecast signals were added to the existing signals (figure below).
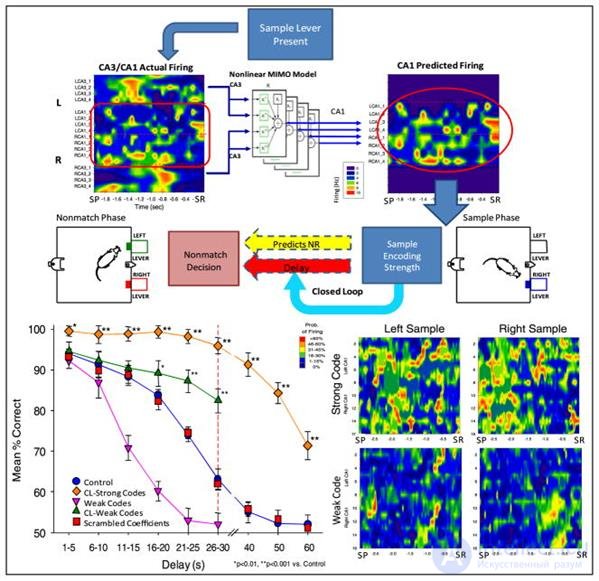 Adding Artificial Signals to Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011
Adding Artificial Signals to Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011 It turned out an interesting result. It turned out that a significant impact on the result is only when the signal is corrected at the stage of sample presentation, that is, when the original image is memorized. The picture of the signals in the remaining moments was irrelevant. Moreover, the stronger the correction signal was, the better was the final result.
The authors of the experiment, explaining the result, proceeded from the idea that the hippocampus is engaged in coding the current information, preparing it for writing to memory. That the pictures of activity observed in the hippocampus carry an informative description of the events taking place. Accordingly, the amplification of the "correct" signal contributes to the improvement of the results of memorization.
In our model, the explanation of the results obtained is somewhat different. It is clear that the hippocampus identifier is important only at the moment of memory formation. Once the memory is created the hippocampus is not required to play it. Strengthening the current hippocampus identifier has a positive effect on the power of memorization, but it is not the result of creating a clearer information picture, but simply highlighting the current memory from the rest of the memory. If a normally working brain thus emphasizes all memories, then the final result will be zero.
The most interesting series of experiments was related to the suppression of the hippocampus own activity. For this, rats were given regular injections into the CA3 region of the hippocampus of the drug MK801. MK801 blocks the normal operation of synaptic transmission using glutamate. It penetrates into the ion channels of NMDA receptors sensitive to glutamate and disrupts their work.
The blockade of the CA3 site has dramatically worsened the expected results. But the supply of artificial signals, restoring the expected pattern of activity, has significantly improved the number of correct answers (figure below).
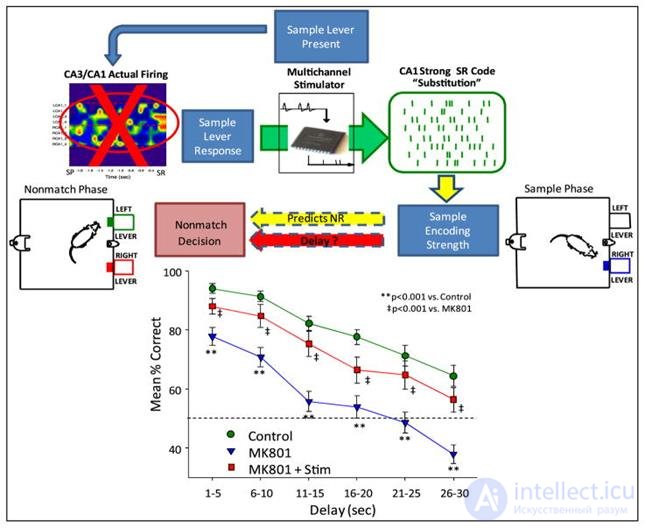 Replacing the natural hippocampal signal with an artificial one (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011)
Replacing the natural hippocampal signal with an artificial one (Theodore W. Berger, Robert E. Hampson, Dong Song, Anushka Goonawardena, Vasilis Z. Marmarelis, Sam A. Deadwyler, 2011) I am not inclined to interpret this result as a wonderful restoration of a neural description of what is happening in the hippocampus, followed by the recording of this description in memory. Especially since matrices with a total of 32 electrodes were used in the experiment, of which about half were involved. Most likely, a random identifier was created, which made possible the formation of memory.
But not everything is so simple with the hippocampus. There are several facts that are quite puzzling when you first meet them. So in 1971, John O'Keefe discovered the place cells in the hippocampus (O'Keefe J., Dostrovsky J., 1971). These cells react like an internal navigator. If a rat is placed in a long corridor, then by the activity of certain cells it will be possible to say exactly in which place it is located. Moreover, the reaction of these cells will not depend on how she got into this place.
In 2005, neurons were found in the hippocampus that encoded a position in space, forming something like a coordinate grid (Hafting T., Fyhn M., Molden S., Moser MB, Moser EI, 2005).
In 2011, it turned out that in the hippocampus there are cells that in a certain way encode time intervals. Their activity forms rhythmic patterns, even if nothing else happens around (Christopher J. MacDonald, Kyle Q. Lepage, Uri T. Eden, Howard Eichenbaum, 2011).
Of all these facts, the conclusion is that the hippocampus is responsible for coding our position, both in time and in space. At the same time, the hippocampus is similar to a soldier, for whom it is quite natural to “dig from the fence to lunchtime”. The hippocampus combines both spatial and chronological navigation, which, by the way, have much in common. Just as our journey in space occurs from a landmark to a landmark, so a journey in time passes from one chronological mark to another.
We have previously argued that the hippocampus forms unique identifiers for memories. How does this feature relate to the fact that the hippocampus tracks our spatio-temporal coordinates? Of course, we can assume that the hippocampus has a dual function, participating in two independent processes. But it is more logical to assume that we are dealing with two sides of the same coin.
How do we identify information at all? Using Google, we compose a query consisting of a set of words that defines the meaning of our search. After receiving the list of results, we can then refine it by entering restrictions on the date or geography.
Creating a library of images, we mark the photos with the date of their creation and the geotagging of the place where they were taken. Then we describe who or what is imprinted on them.
Placing files on your computer, we specify the time of creation and the path to the file. The file name, we briefly describe its content.
It turns out that identifying the most different things, we use, on the one hand, signs describing their meaning, and, on the other hand, coordinates in space and time, as universal signs of any phenomena. Such identification subsequently turns out to be quite convenient and useful not only as a unique label, but also as a search tool. It can be assumed that nature went the same way.
That is, creating a unique event identifier, it makes sense for the hippocampus not only to generate a random code and distribute it throughout the cortex, but to lay out the space-time features in this code as the most suitable for all occasions. Such a spatial-temporal description itself is quite unique, as a combination of various factors. It is enough to add to it a certain small random appendage and the resulting identifier will uniquely indicate a specific event.
Such identification is much more powerful in its capabilities than just a unique key for each memory. There is the possibility of a variety of additional associations. But most importantly, it is possible to encode complex descriptions that take into account how events develop in both time and space. But talk about this will be a little later.
If we assume that the hippocampus identifier actually encodes the space-time coordinates, it turns out that the hippocampus, firstly, has the ability to interact with the cortex in such a way as to be able to learn to recognize similar positions in space and time. Second, in order to spread their identifiers throughout the entire cortex, the hippocampus must form them from a finite number of unique fragments. This is due to the fact that the crust must be pre-trained to distribute the entire "alphabet" used by the hippocampus. New identifiers should not be formed "from scratch", but as combinations of already known elements. In principle, these requirements are not difficult, it corresponds to the way in phrases in natural language all phrases are built from a finite set of letters.

Comments
To leave a comment
Logic of thinking
Terms: Logic of thinking