
This series of articles describes a wave model of the brain that is seriously different from traditional models. I strongly recommend that those who have just joined begin reading from the first part.
Let's try to present the information capacity of the brain. Since the memory is based on neurons, for the beginning we will clarify how many neurons we are dealing with. The most accurate estimate for today is Suzanne Herculano-Huzel (Frederico AC Azevedo, Ludmila RB Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata EL Ferretti, Renata EP Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009 ). According to this estimate, the cortex accounts for 82% of the total brain mass and 19% of the total number of neurons (figure below). According to this estimate, the cortex consists of about 16 billion neurons (1.6x10
10 ).
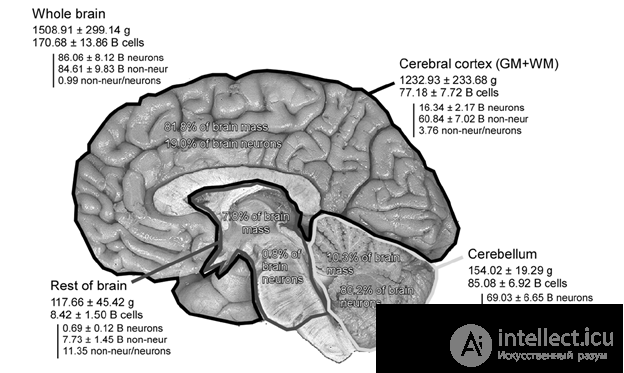 The number of neurons in different parts of the brain (in billions) (Frederico AC Azevedo, Ludmila RB Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata EL Ferretti, Renata EP Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009)
The number of neurons in different parts of the brain (in billions) (Frederico AC Azevedo, Ludmila RB Carvalho, Lea T. Grinberg, José Marcelo Farfel, Renata EL Ferretti, Renata EP Leite, Wilson Jacob Filho, Roberto Lent, Suzana Herculano-Houzel, 2009) In models that use the plasticity of synapses to explain the nature of memory, memory capacity is considered quite simple. The characteristic number of synapses for a neuron is 10,000 (10
4 ). If we assume that one synapse stores one bit of information, then we get 1.6x10
10 x 10
4 = 1.6x10
14 bits or approximately 2x10
13 bytes. This is about 20,000 gigabytes or 20 terabytes. The volume is quite modest even by today's standards.
If we assume that the active life of a person is about 80 years old, of which he spends one third in a dream, then it will be 80x365x (24-8) x60x60 = 1681920000 (1.7x10
9 ) seconds. It turns out that if you do not take into account the organization of memory, but simply fill it in during life, then you can allow the write speed of 2x10
13 / 1.7x10
9 = 1.18x10
4 bytes / second or about 10 kbytes / second.
In our model, an estimate of the amount of memory looks a little different. Evaluation of synaptic memory does not change, the same 20 terabytes. But this is only a memory that stores images of neuron detectors. The main volumes are in extrasynaptic receptive clusters. If one assesses that one extra-synaptic image is formed from signals of about 100 synapses, then the information capacity of one element can be estimated as 100 bits. Based on the estimate that the surface of the neuron can contain about 100,000 receptive clusters, we get a memory capacity of 1.6x10
10 x10
5 x100 = 1.6 x10
17 bits or 2x10
16 bytes. What is 20,000 terabytes or 20 petabytes.
If we assume that half of this memory is spent on the ability of the cortex to propagate identifier waves, then the amount of event memory can be estimated at 10 petabytes. This volume causes a certain respect. Based on it, the speed of recording information during continuous memorization can be 10
16 /1.7x10
9 = 5.88x10
6 bytes / second or about 6 megabytes / second. If we assume that the fixation of engrams occurs several times per second, then we can estimate the information capacity of one engram in the region of 1 megabyte.
Potentially, our memory is able to store a fairly detailed description of everything that happens to us in life. The totality of memory is confirmed by the presence of people-phenomena that truly "remember everything." Alexander Luria, as a result of many years of observation, has documented one example of such an infinite memory:
“It turned out that S.'s memory does not have clear boundaries not only in its volume, but also in the strength of holding traces. Experiments have shown that with success — and without noticeable labor — he can reproduce any long series of words given to him a week, a month, a year, many years ago. Some of these experiments, which invariably ended in success, were carried out 15 to 16 years later (!) After the initial memorization of a series and without any warning. In such cases, Sh. Sat down, closed his eyes, paused, and then said: “Yes, yes ... it was in your apartment ... you sat at the table, and I was on the rocking chair ... you were in a gray suit and looked at me like that ... now ... I see that you told me ... "- and then followed an unmistakable reproduction of the series read" (Luria, 1968).
But no matter how infinite our memory is, the fact is well known that we recall recent events much easier and in more detail. As early as 1885, the forgetting curve (Ebbinghaus, 1885) was obtained by psychologist Hermann Ebbinghaus (figure below).
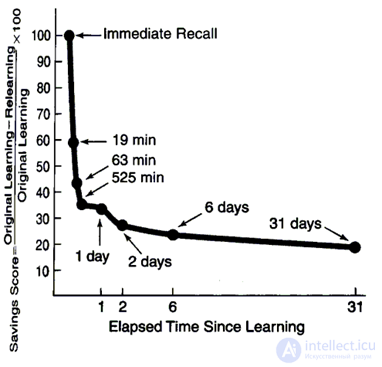 Forgetting curve
Forgetting curve The sharpest forgetting is observed in the first minutes, then the process of forgetting slows down. After 30 days, the memory enters a stable phase when further forgetting does not occur. In his experiments, Ebbinghaus used sets of meaningless three-letter words that he memorized. Memorizing is somewhat different from simple perception, that is, the forgetting curve for a phrase that was heard once or a seen image will be somewhat different.
To remember something more strongly allows repetition. Repeating the material, we do not just refresh our memories and begin a new cycle of forgetting. After the repetition, the asymptote, on which the forgetting curve goes, rises significantly higher in comparison with its previous level (figure below). Each new repetition increases the chances that, after a long time, we will be able to recall everything relatively unmistakably.
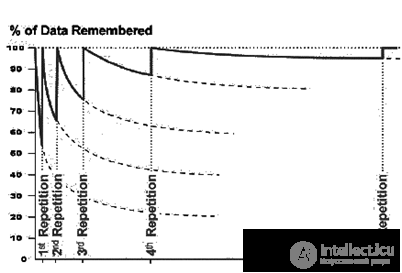 Curve forgetting with repetitions
Curve forgetting with repetitions It is important to note that repetition does not necessarily have to be reduced to one more presentation of a memorized image. A memory that accurately reproduces a memorized event is also a repetition. That is, the more often we remember something, the stronger it bumps into our memory.
The traditional interpretation of such properties of memory is based on the idea of transferring memories from short-term memory to long-term memory. At the same time, often based on computer analogies of data storage, they imply the transfer of information traces from one part of the brain to another. So it is assumed that the hippocampus, working like computer’s memory, accumulates current memories and then moves them to the cortex, in the same way as saving computer data on long-term media.
The process of transition of memories from the short-term to the long-term form is called memory consolidation. This is an objective and fairly well-studied process. Besides the fact that we can observe changes in memory on our own experience, there are also direct confirmations of the presence of consolidation. So it is known that head injuries, electric shock, cramp violate the normal course of memory formation. They cause retrograde amnesia, when memories related to a certain period of time preceding the failure are lost.
A similar effect of memory loss is obtained when substances that block the expression of certain genes that produce proteins involved in memory work are introduced into the brain.
In addition, the effect of retrograde amnesia is familiar to many for significant memory loss after excessive drinking. A certain critical dose of alcohol consumed so much violates the process of consolidation, that part of the memories is completely erased, creating an irreparable failure in memory. In this case, there is no loss of the ability to memorize, as with disturbances in the work of the hippocampus, namely, erasing the already received memories, or rather not translating them into a long-term storage phase. A patient with Korsakov syndrome loses memory as soon as his attention switches. A person who has drunk a critical dose still remembers recent events, and in many cases can maintain a related conversation. However, in the morning all the memories, starting from a certain point, are erased.
To explain the effect of consolidation, it is not necessary to close on a computer analogy associated with the physical movement of data. In our model, consolidation and its meaning can be described as follows. As we said earlier, extrasynaptic metabotropic receptive clusters have a hysteresis effect. This means that if a neuron generates a spike, then a jump in the membrane potential under the fulfillment of a number of conditions described by us earlier can serve as a reason for the transition of the receptive cluster to a steady state. The steady state is either a state in which the end portions of the receptors are pushed out of the membrane and the receptor becomes sensitive to a specific signal, or the state of their drowning in which the receptor, on the contrary, loses sensitivity. Such a change occurs with receptive clusters for which the condition of interference of mediators from the synapses surrounding them has been fulfilled, that is, they have tuned in to a particular image.
The transition of receptive clusters to this state occurs simultaneously in a huge number of neurons that are active at the current time. Which means that this current moment is remembered with extreme redundancy and granularity. This leads to the fact that, all other things being equal, current events have a much higher probability of being recalled as compared with events more distant in time.
Such a hyper-detailed memory can be called operational or short-term memory, but do not confuse it with the current induced and wave activity of cortical neurons. The activity of neurons forms a description of what is happening, while the short-term memory stores pictures of such relatively recent states. Moreover, short-term memory is no different from the rest of the memory, except for the fact that fresh memories are more detailed, since they use a significantly larger number of receptive clusters than other engrams.
The length of the super-fast memory is seconds. Then most of the extrasynaptic receptors return to their original state, gaining readiness to participate in new memorization acts. But a significant part of receptive clusters for one reason or another remains in a state of fixation of the image. They move into the next stable phase, which allows them to exist further and accordingly store a fragment of their memories.
It can be assumed that each memory goes through several stages of consolidation, losing part of its receptive clusters at each stage. The easiest forgetting algorithm is probabilistic. By adjusting the probability of the next consolidation stage being passed by the receptive cluster, it is possible to obtain the required volume for each memory by the time of its transition to long-term memory.
The consolidation phase is accompanied by the synthesis of advising proteins, which determine the overall course of all processes. The course of consolidation for metabotropic and ionotropic receptors has its own characteristics, but is quite similar ideologically. Immediately after training, the expression of immediate "early genes" begins in the neuron nucleus. These genes encode transcription factors. Transcription factors trigger the transcription of “late genes”. The approximate schedule of such a procedure is shown in the figure below.
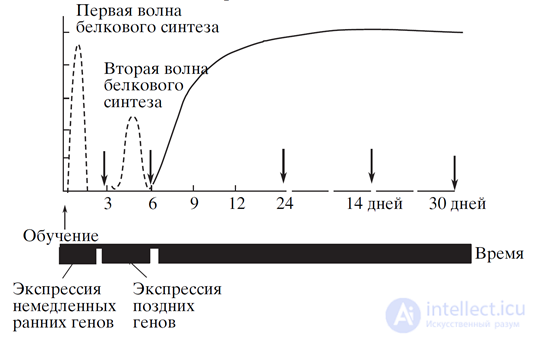 Expression of the "early" and "late" genes (E.V. Muravyeva, K.V. Anokhin, 2006).
Expression of the "early" and "late" genes (E.V. Muravyeva, K.V. Anokhin, 2006). Various proteins are known, in relation to which their participation in the process of memory formation is proved. Arc, CPEB, Kibra - this is only a small part of them, although the most frequently mentioned. We must remember that information leaves two types of traces on a neuron. One is associated with a change in the sensitivity of its synapses, the other with the fixation of changes in extrasynaptic receptors. The first affects the properties of the neuron as a detector. The second provides the ability of neurons to propagate waves of identifiers, store event memory, form associativity, and the like. How and in what sequence the protein regulation of these processes takes place is not a simple question and has not yet had a generally accepted answer, since there is not yet a generally accepted memory model. Therefore, in our model we will focus on the general ideology of the processes, unfortunately, it is not the name of the opportunity to conclusively describe their protein regulation.
Consolidation actually sets the timer, which counts the life stage of the elements of the memory and regulates the probability of their transition to the next phase. It can be assumed that the repetition of experience or, equivalently, the recollection of it, alter the course of consolidation, increasing the chances of receptive clusters to go into the phase of long-term memory. That is, the more repetitions or the more often the memories, the more significant the trace leaves the event. And the higher the probability of its recovery with the corresponding information request.
Since events that are emotionally significant cause us to recall them more often than neutral events, this may be one of the reasons why emotionally colored events form stronger and more stable traces in our memory.
But the memories of the experiences are not as simple as they seem. Psychologists have long known the phenomenon of memory transformation. It was first shown in 1932 by Frederick Bartlett (Bartlett, 1932). He found that if a person, recalling a certain event, adds any details, the original memory is replaced by a new one, and the changed details become part of this new memory. And the person is not aware of these changes, and believes that everything was exactly as he remembers now. Such a change in memories is called memory reconsolidation.
Memory re-consolidation fits well with our model. The memory of an event is accompanied by the restoration of its unique identifier. If for any reason the original memory is distorted, the current information image is remembered with the same identifier as the main memory, since this identifier is present in the space of the cortex. This means that the transformation phenomenon described by Barlett inevitably happens.
There are very interesting experiments that show that reconsolidation can be accompanied not only by transformation, but also by erasing memory. But since these experiences are more related to the behavior than to the event memory, we will consider them later in the appropriate section.
The general ideology of event memory can be described as follows. The incoming information creates fresh memories with great redundancy and detail. This makes them extremely accessible for online access. Such a memory does not live long. She memory is subjected to stages of consolidation, each of which reduces the number of receptive clusters that form engrams. As a result of all stages of consolidation, which apparently lasts for about a month, engrams acquire a stable volume and pass into the phase of long-term storage.
Earlier, we estimated the order of the volume of brain event memory at 10 petabytes. From where the gypsy method derived an estimate of the size of an engram of 1 megabyte. This assessment may be valid for old, past all stages of consolidation, memories. Most of the information with which we operate, refers to the current day. Deployed redundant storage is provided for this information. It can be assumed that such actual memory takes up a substantial amount of the total brain memory. If I were the designer of the human brain, then I would empirically take half of all available memory to fresh memories, and define the remaining half as old memories.
With this empirical assumption, the speed of recording information by the brain can be estimated as 5x10
15 / (16x60x60), which is about 10
11 bytes or 100 gigabytes per second. Of course, this is much more than the entire amount of information coming from all the senses together. But the conversation is not about the input volume, but about the detailed description, consisting of the set of descriptions that arise on each of the zones of the cortex.
In this estimation, the initial size of one engram will be about 10 gigabytes. Actually, the process of memory consolidation is a gradual reduction of this extremely excessive volume to the level of about 1 megabyte.
However, whatever the ratio of the volumes of new and old memory really was, nature had to optimize it so as to prevent unreasonable redundancy of idle volumes. This means that, given the extremely high speed of recording current information, there may be a moment of overflow of all available memory. It can be assumed that drowsiness - this is the state when the information resources of the brain are suited to their filling, and memory clearing is required.
In principle, operational memory consolidation occurs constantly, continuously compressing the volume of engrams, the brain provides space for new memories. But it can be assumed that operational consolidation has several disadvantages. So in our model, neuron-detectors form patterns that correspond to concepts. Each concept is formed by a set of patterns located in different places of the cortex. For a pattern to trigger a wave, an essential part of the pattern's neurons must be activated. This means that if we use probabilistic consolidation, simply removing some of the receptive clusters, we will gradually destroy all the engrams to such a level that none of the patterns will be able to radiate the wave associated with this memory. To avoid this, it is wise to use selective consolidation.
Selective consolidation assumes that we selectively determine on which patterns it is worth leaving clear traces of memories, and on which it is worth removing them completely. Then, with the same volume of an engram and preservation of its distributed nature, it will be possible to avoid low density of engrams on separate patterns.It can be assumed that it is more advantageous to use those patterns that preserve on the cortex most closely correspond to the activity associated with the general context of the memory to save engrams. But the procedure for determining such patterns requires repeated technical reproduction of the memorized events. However, such reproduction is incompatible with the normal functioning of the brain, since, proceeding simultaneously with the current perception, it will be perceived by the brain as a hallucination. It is appropriate to assume that optimal consolidation associated with the removal of most of the information accumulated during the day is not possible or difficult during normal brain work. Then it turns out that sleep is precisely the condition that is necessary for the brain to perform procedures of this kind.In addition to the fact that the brain accumulates event memories, at the same time, neurons are trained as detectors. Both are based on changes in the structure of neural receptors. Changes in receptors are driven by the expression products of certain genes. Gene expression products are proteins. Some of them are directly involved in cellular processes, some are transcription factors that regulate the subsequent expression of genes. By changing the concentration of the corresponding proteins, one can judge the course of the processes associated with both types of consolidation.Protein concentration can be seen and assessed through staining of the brain tissue with a dye that selectively reacts to the desired protein. Similar observations have shown that the most significant changes occur during sleep (Chiara Cirelli, Giulio Tononi, 1998) (Cirelli, 2002) (figures below).  Arc protein distribution in rat parietal cortex after three hours of sleep (S) and after three hours of spontaneous wakefulness (W) (Cirelli, 2002)
Arc protein distribution in rat parietal cortex after three hours of sleep (S) and after three hours of spontaneous wakefulness (W) (Cirelli, 2002)  Distribution of P-CREB transcription factor in coronal areas of rat parietal cortex after three hours of sleep (S) and in the case of sleep deprivation for three hours (SD) (Cirelli, 2002) Memory consolidation in a dream occurs in several stages. In a healthy person, sleep begins with the first stage of slow sleep, which lasts 5-10 minutes. Then comes the second stage, which lasts about 20 minutes. Another 30-45 minutes are in the periods of the third and fourth stages. After that, the sleeper again returns to the second stage of slow sleep, after which the first episode of REM sleep occurs, which has a short duration of about 5 minutes. During REM sleep, the eyeballs very often and periodically make quick movements under closed eyelids. If at this time to wake the sleeper, then in 90% of cases you can hear the story of a bright dream. This whole sequence is called a loop. The first cycle has a duration of 90-100 minutes. Then the cycles are repeated,at the same time the share of slow sleep decreases and the share of fast sleep gradually increases, the last episode of which in some cases can reach 1 hour. On average, with full healthy sleep, there are five complete cycles.Sleep stages can be explained by the need to process various types of information. The algorithms for consolidating event memory, wave memory associated with the propagation of identifiers, and memory associated with the plasticity of synapses can vary greatly.In our assumptions, the cyclical nature of sleep is well explained. Full consolidation, as everyone knows from personal experience, requires about eight hours. But circumstances can make us wake up earlier. If the eight-hour sleep consisted of one cycle, divided into several phases, the premature interruption of sleep would lead to the fact that the initial phases of sleep would have completed completely, while the later ones did not even begin. That is, some of the information would remain completely unprocessed, while completely exhausting the corresponding brain resource for the previous day. That is, the brain, in fact, would be inoperable. In the case of a premature awakening, several cycles allow at least partially performing each of the consolidation procedures and bringing the brain to an adequate state. But the need for full consolidation does not disappear,and subsequently, the brain will have to compensate for lost cycles with a longer sleep, which is usually called “sleep.”It seems that the consolidation of memory is associated with the phases of slow sleep. As it completes, these phases become shorter, giving way to fast sleep. This explains the fact that with the full completion of consolidation, the final stage of sleep consists solely of the prolonged phase of REM sleep. Fast sleep - these are the very dreams that form bizarre fantasies, only remotely related to real experience. The pattern noted for dreams even by Sigmund Freud, says that in a dream appear the phenomena most disturbing to us, important for us, concentrating the essence of our torments and experiences (Freud, 1900). In most cases, dreams are very quickly forgotten. We see dreams every night, but the swiftness with which we forget them makes many believe that they do not see dreams at all.In our model, erasing dreams is implemented quite simply. To do this, it is enough to make sure that traces of the hippocampus identifiers are not fixed for a long time, but are subject to rapid dissolution. This will lead to the fact that we will permanently forget about dreams. But if, on waking up, we have time to recall a recent dream, then a new traditional memory will arise that will preserve the events of sleep. It can be assumed that for dreams, erasing traces of hippocampal identifiers is not accompanied by removal of associative traces. Then the meaning of dreams can be explained by the strengthening of the system of associations for the most significant and important concepts for us. Which in turn contributes to the subsequent perception of the world in such a way that these concepts will be most emphasized in the system of describing new events.It is well known to all that “the morning of the evening is wiser”. But what is the essence of this wisdom? Sometimes morning insights are explained by the fact that de, they say, the brain at night solves daily tasks. This explanation is prompted, firstly, by the fact that the overnight activity of the brain recorded in many studies strongly resembles the activity experienced during the day, and, secondly, the consonance of dreams with themes that disturb us in reality. But it is more appropriate to assume that the brain does not solve daytime tasks at night, it is working to optimize the system of concepts and memories. In the morning this optimization makes it much easier to find explanations yesterday for still incomprehensible questions. To properly ask a question, you need to know most of the answer. That jobwhat the brain does at night — this is the preparation of the largest part of the answer — the possibility of approaching the issue with the “right” set of internal concepts and getting rid of unnecessary detail of unnecessary memories. That is, if everything said now about sleep seems too vague to you, then try to return to this tomorrow.It makes no sense to regret the time spent in a dream. Sleep is a great opportunity to “get smarter” without making any effort. The only thing that is necessary is to pre-load the head with a sufficient amount of fresh information. Moreover, this information can be not only new facts, but also old knowledge, newly recalled or deliberate. Any of our thoughts leave a trace of fresh memories on the bark. In the dream, there is a consolidation of new reflections and old experience, which allows deeper generalizations to arise. This means that daytime thinking on any question makes sense, even if they do not lead to an immediate answer. These reflections create fresh memories related to the topic that interests us, which at night will give the brain the opportunity to work on optimizing the entire system of our concepts.It may be recalled that small children, in whom the organization of the cortex is most active, in general, spend most of their time in a dream.The ideas of consolidation can explain not only the sleep of infants, but also the conservatism of the elderly. By conservatism, we understand the rejection of new ideas and skills, provided that we maintain normal event-based memory. In traditional approaches, this causes a certain contradiction. It would seem that the ability to memorize new facts should ensure the ability to learn new principles and learn new skills. But everything falls into place if we remember that there is an organization of neuron detectors that forms a set of concepts of the brain, but there is an event memory described and stored on the basis of these concepts. Daily consolidation of event memory clears up, apparently, more than half of the available volume. This means that, regardless of age, event memory can be strong if organic disturbances do not interfere with this.But with neuron detectors everything is more complicated. It seems that with age, the number of neurons that are determined with their specialization is becoming more and more, and there are fewer and fewer opportunities for the formation of new concepts. And then we come to a situation that we can remember everything, but only in those concepts that we have. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism.which we exist. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism.which we exist. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism.
Distribution of P-CREB transcription factor in coronal areas of rat parietal cortex after three hours of sleep (S) and in the case of sleep deprivation for three hours (SD) (Cirelli, 2002) Memory consolidation in a dream occurs in several stages. In a healthy person, sleep begins with the first stage of slow sleep, which lasts 5-10 minutes. Then comes the second stage, which lasts about 20 minutes. Another 30-45 minutes are in the periods of the third and fourth stages. After that, the sleeper again returns to the second stage of slow sleep, after which the first episode of REM sleep occurs, which has a short duration of about 5 minutes. During REM sleep, the eyeballs very often and periodically make quick movements under closed eyelids. If at this time to wake the sleeper, then in 90% of cases you can hear the story of a bright dream. This whole sequence is called a loop. The first cycle has a duration of 90-100 minutes. Then the cycles are repeated,at the same time the share of slow sleep decreases and the share of fast sleep gradually increases, the last episode of which in some cases can reach 1 hour. On average, with full healthy sleep, there are five complete cycles.Sleep stages can be explained by the need to process various types of information. The algorithms for consolidating event memory, wave memory associated with the propagation of identifiers, and memory associated with the plasticity of synapses can vary greatly.In our assumptions, the cyclical nature of sleep is well explained. Full consolidation, as everyone knows from personal experience, requires about eight hours. But circumstances can make us wake up earlier. If the eight-hour sleep consisted of one cycle, divided into several phases, the premature interruption of sleep would lead to the fact that the initial phases of sleep would have completed completely, while the later ones did not even begin. That is, some of the information would remain completely unprocessed, while completely exhausting the corresponding brain resource for the previous day. That is, the brain, in fact, would be inoperable. In the case of a premature awakening, several cycles allow at least partially performing each of the consolidation procedures and bringing the brain to an adequate state. But the need for full consolidation does not disappear,and subsequently, the brain will have to compensate for lost cycles with a longer sleep, which is usually called “sleep.”It seems that the consolidation of memory is associated with the phases of slow sleep. As it completes, these phases become shorter, giving way to fast sleep. This explains the fact that with the full completion of consolidation, the final stage of sleep consists solely of the prolonged phase of REM sleep. Fast sleep - these are the very dreams that form bizarre fantasies, only remotely related to real experience. The pattern noted for dreams even by Sigmund Freud, says that in a dream appear the phenomena most disturbing to us, important for us, concentrating the essence of our torments and experiences (Freud, 1900). In most cases, dreams are very quickly forgotten. We see dreams every night, but the swiftness with which we forget them makes many believe that they do not see dreams at all.In our model, erasing dreams is implemented quite simply. To do this, it is enough to make sure that traces of the hippocampus identifiers are not fixed for a long time, but are subject to rapid dissolution. This will lead to the fact that we will permanently forget about dreams. But if, on waking up, we have time to recall a recent dream, then a new traditional memory will arise that will preserve the events of sleep. It can be assumed that for dreams, erasing traces of hippocampal identifiers is not accompanied by removal of associative traces. Then the meaning of dreams can be explained by the strengthening of the system of associations for the most significant and important concepts for us. Which in turn contributes to the subsequent perception of the world in such a way that these concepts will be most emphasized in the system of describing new events.It is well known to all that “the morning of the evening is wiser”. But what is the essence of this wisdom? Sometimes morning insights are explained by the fact that de, they say, the brain at night solves daily tasks. This explanation is prompted, firstly, by the fact that the overnight activity of the brain recorded in many studies strongly resembles the activity experienced during the day, and, secondly, the consonance of dreams with themes that disturb us in reality. But it is more appropriate to assume that the brain does not solve daytime tasks at night, it is working to optimize the system of concepts and memories. In the morning this optimization makes it much easier to find explanations yesterday for still incomprehensible questions. To properly ask a question, you need to know most of the answer. That jobwhat the brain does at night — this is the preparation of the largest part of the answer — the possibility of approaching the issue with the “right” set of internal concepts and getting rid of unnecessary detail of unnecessary memories. That is, if everything said now about sleep seems too vague to you, then try to return to this tomorrow.It makes no sense to regret the time spent in a dream. Sleep is a great opportunity to “get smarter” without making any effort. The only thing that is necessary is to pre-load the head with a sufficient amount of fresh information. Moreover, this information can be not only new facts, but also old knowledge, newly recalled or deliberate. Any of our thoughts leave a trace of fresh memories on the bark. In the dream, there is a consolidation of new reflections and old experience, which allows deeper generalizations to arise. This means that daytime thinking on any question makes sense, even if they do not lead to an immediate answer. These reflections create fresh memories related to the topic that interests us, which at night will give the brain the opportunity to work on optimizing the entire system of our concepts.It may be recalled that small children, in whom the organization of the cortex is most active, in general, spend most of their time in a dream.The ideas of consolidation can explain not only the sleep of infants, but also the conservatism of the elderly. By conservatism, we understand the rejection of new ideas and skills, provided that we maintain normal event-based memory. In traditional approaches, this causes a certain contradiction. It would seem that the ability to memorize new facts should ensure the ability to learn new principles and learn new skills. But everything falls into place if we remember that there is an organization of neuron detectors that forms a set of concepts of the brain, but there is an event memory described and stored on the basis of these concepts. Daily consolidation of event memory clears up, apparently, more than half of the available volume. This means that, regardless of age, event memory can be strong if organic disturbances do not interfere with this.But with neuron detectors everything is more complicated. It seems that with age, the number of neurons that are determined with their specialization is becoming more and more, and there are fewer and fewer opportunities for the formation of new concepts. And then we come to a situation that we can remember everything, but only in those concepts that we have. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism.which we exist. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism.which we exist. But to form new concepts, we can hardly or not at all. Accordingly, everything fundamentally new causes perception difficulties and natural rejection, which is called senile conservatism. 
Comments
To leave a comment
Logic of thinking
Terms: Logic of thinking