Lecture
The lowest level of perceptual development of the psyche
Perceptual psyche is the highest stage of development of mental reflection. Recall that this stage is characterized, according to Leontiev, by a change in the structure of the activity - by highlighting the content of the activity relating to the conditions in which the object of activity is given in the environment (operation). For this reason, we meet here with genuine skills and perceptions. Subject components of the environment are already reflected as integral units, while in the elementary sensory psyche there was a reflection of only some of their properties or the sum of the latter. Object perception necessarily implies a certain degree of generalization, sensual representations appear.
Of course, the perceptual psyche, peculiar to a huge number of animals standing on different levels of evolutionary development, reveals great differences in their specific manifestations. Therefore, it is also necessary at this stage of development of the psyche to single out individual levels, of which the lowest will be considered here.
At this level, there are primarily higher invertebrates - cephalopods and arthropods (at least higher). We will give the characteristics of the considered level of mental development using the example of insects, a central class of the type of arthropods. (In addition to insects, arthropods include crustaceans, arachnids, millipedes, and some small groups.) Insects are the most numerous class of animals, both in the number of species and in the number of individuals. Insects live everywhere on land (in all climatic zones) - both on the surface and in the soil, in all fresh water bodies, as well as in the air, rising to an altitude of two kilometers.
Movements
The huge variety of “ecological niches” occupied by insects and other arthropods, naturally led to the development of very different forms of adaptation in the field of the structure and behavior of these animals. The most significant and characteristic of the evolution of arthropods, especially insects, was the conquest of the new habitat - land. Arthropods are the first terrestrial animals in Earth history. The transition to the land was associated with the development of special organs of movement - the limbs in the form of complex arms consisting of separate joints connected by joints, according to which arthropods got their name. Such complex levers (their analogs are the limbs of terrestrial vertebrates) are best used to move along a solid substrate by pushing it away. Arthropods are able to move over land in almost all possible ways (crawling, walking, running on both horizontal and vertical surfaces, jumping, movement in the depth of the ground). In water, arthropods swim, dive, crawl, and even run on its surface.
Insects, moreover, in connection with the transition to life in the air environment, a completely new way of movement appeared for the first time - flying with the help of wings. Insects are able to develop a very high speed in flight: for some dragonflies it reaches 90 kilometers per hour, and even 122 kilometers per hour for a male gadfly. The frequency of wing flaps in many insects is truly amazing. So, the moth makes only 9 beats per second, the same bee makes 190 bees, and the housefly makes 330 but in some flies and hymenoptera, the speed of the wings vibrates up to a thousand beats per second. No other animals have observed such rapid movements and, accordingly, the frequency of muscle contractions.
Accordingly, the wing musculature of insects is distinguished by an exceptionally complex device: it consists of a multitude of differently located muscle bundles, partially acting as antagonists. Other departments of the muscular system are also finding it more difficult.
In general, insects (apart from the wing muscles of insects) distinguish the following sections of the muscular system: the muscles of the body, which control its movements: the muscles of the limbs, through which the whole limb moves in relation to the body; muscles of individual limb segments, muscles of internal organs.
As can be seen, the musculature of insects is distinguished by a high degree of differentiation and specialization. In addition, the vast majority of their muscles - striated. The number of individual muscles is exceptionally large - one and a half thousand and more. Muscle strength is also impressive: insects are capable of carrying objects, sometimes a hundred times their own weight. In general, as Dogel pointed out, in arthropods the muscular system of invertebrates reaches the “apogee of its development” both in morphological and physiological terms.
central nervous system
In accordance with the complex and highly differentiated organization of the musculoskeletal system, there is also a complex structure of the central nervous system of insects, which, however, we can only characterize here in very general terms.
Like the ringed worms, insects are metamerically located ganglia, the base of the entire nervous system, interconnected by nerve trunks. These trunks converge in the majority of insects so that they form (at least outwardly) one unpaired nervous chain. Simultaneously, the merging of successive metameric ganglia takes place.
In connection with a clearly pronounced cephalization in the head section, the fused ganglia formed a powerful brain in the form of a complexly arranged epipharyngeal node, in which there are three sections: the protocerebrum, deitocerebrum and tritocerebrum. The anterior section, the protocerebrum, is more developed than the rest. Particularly noteworthy are the so-called mushroom (or stalk-like) bodies, associative and coordinating centers. They are most highly developed in insects with particularly complex behaviors — bees, primarily working individuals, and ants. A pair of very large and complexly arranged visual lobes innervating complex eyes is also associated with the protocerebrum. Daytocerebrum (middle part of the brain) innervates the antennae, tritosecerebrum innervates the mouth area and, moreover, is closely connected with the sympathetic nervous system. In general, insects are characterized by a high concentration of nerve elements. This concentration increases the ability to control individual organ systems and raises the overall morphophysiological level of the body as an integral unit.
The nervous system, and in particular the brain, is arranged in different arthropods, however, it is very different. Significant differences are found between lower and higher representatives, even within classes, and sometimes units. For example, the ratio of the size of the brain to the volume of the body is 1: 3290 in the May beetle, 1: 280 in the ant, and 1: 174 in the bee.
The nervous system of cephalopod mollusks is at a very high level of development, radically different from that of other mollusks. This is especially true of their very complex brain. Attention is drawn to the exceptional concentration of the brain ganglia: the ganglia have merged together and form a total perioesophageal mass. The bases of the optic nerves form enormous swellings — the optic ganglia, which by their size often exceed the cerebral nodes themselves.
Nerves extending from the brain to effectors reach very high power — up to a millimeter and larger in diameter, which is 5–10 times the thickness of the nerves of most vertebrates! This provides a much higher rate of nerve impulses than the lower mollusks. Accordingly, the enormous size of the eye is exceptionally developed and the optic nerve. The outstanding Soviet histologist A. A. Zavarzin showed a great similarity of the fine histological structure of the optical centers of cephalopods and vertebrates.
Thus, we see that the profound differences between the elementary sensory and perceptual psyche are embodied in the structure of the nervous system already at the lowest level of the perceptual psyche.
Sensory abilities, taxis
The specific conditions in which insects live, the far-reaching adaptation of motor activity with an abundance and variety of qualitatively very different agents of the environment that govern their behavior — all this caused the appearance in these animals of many, and moreover, peculiarly arranged, sense organs.
Vision plays a big role in the life of insects. In different numbers, different species have simple single eyes. But the main organ of vision is a kind of complex eye, called facet, because its surface consists of separate, clearly limited areas - facets. Each facet corresponds to a separate eye - ommatidium, isolated from its neighbors by a pigment pad and functioning independently of them. The total outer cover of the eye forms a thickening above the ommatidia, referred to as the cornea or horn lens and playing the role of the eye dioptric apparatus. Each ommatidium contains 7–8 visual cells, the axons of which terminate in the optical ganglia of the brain.
Thanks to such a device of complex eyes, “mosaic” images of perceived subject components of the environment are transmitted to the insect brain. Of course, this does not mean that the insect really sees the world around it in the form of a mosaic. In the brain, visual stimuli coming from individual ommatidia are synthesized into a complete picture, as is the case with vertebrates. Yet it is very difficult for us to imagine how an insect actually sees.
Even in the last century, attempts were made to photograph, through photographic eyes, the clarity and range of insects and crayfish. Later, in particular in the 60s of our century, such experiments were repeated, but it is hardly possible in this way to solve the problem, because, as we know, it is not the eye that sees, but the brain. Insects are obviously quite short-sighted: clear vision, for example, of the urticaria butterfly is possible only at a distance of about 2–3 centimeters. From objects located further (starting from 5–7 centimeters), insects receive only vague images, but, obviously, this is enough for their general orientation in space.
At a distance, the bee is able to see only objects a hundred times larger than is necessary for human vision. She sees a single flower only when it comes close to it. All these features of insect view are quite consistent with the microlandscape in which these animals live.
Visual acuity (resolution, i.e., the ability to see two close points) in insects, all other things being equal, in particular, on the number of ommatidia, which, however, fluctuates greatly even within orders. Thus, in a detachment of beetles, this number ranges from three or four dozen to several tens of thousands. In a bee, each eye consists of approximately four thousand facets. The smaller the facets, the weaker the light in them, but the more detailed the vision. This is especially important for the objective view. In addition, with a more prominent eye, the ommatidia axis diverges more strongly, and as a result, the field of view expands. At the same time, the point seen by one facet narrows, which also increases visual acuity.
In the best way, insects see moving objects or fixed objects during their own movement. A bee flying over a meadow, for example, sees flowers swaying in the wind, or an image of a still flower glides over its faceted eye. In any case, this mobile (vague) image causes it to go down and examine the plant from the minimum, i.e., optimal for the bee's vision, distance.
As for color vision, their visible spectrum is wider than that of a person, since its boundary lies in the ultraviolet sector. The controversial question about the ability of insects to perceive red color is now, obviously, resolved in a positive sense (with the exception of the honeybee and some other insects). The Soviet entomologist G. A. Mazokhin-Porshnyakov believes that in general, color vision in adult insects is a universal phenomenon. According to him, the colors differ even in species whose active life takes place at night.
So far, we have dealt only with the reaction of insects to light, which plays a very large role in their lives. No less, if not more important for them smells. In the natural behavior of insects, stimuli of various modalities always act as a complex. Thus, the already mentioned wasps of the genus Philanthus hunt for bees, guided by sight from a distance of 30 centimeters, but identify them by smell only from a distance of 15–5 centimeters. During a visual search, the wasp does not react at all to the smell of the bee.
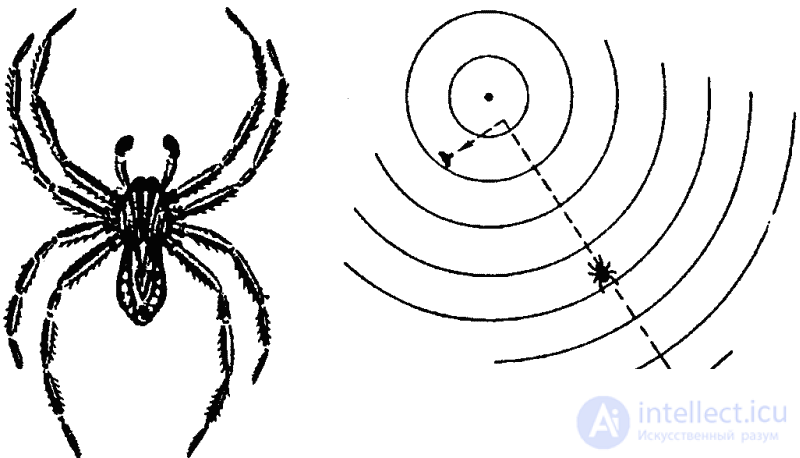
The situation is similar for other arthropods. Water spiders of the genus Pirata orient themselves primarily by the vibration of the surface of the water, and in the immediate vicinity of the victim, by sight (Fig. 36). Other spiders learn about the insect entering their network by perceiving the vibrations of the web. But the victim is examined in three ways at once: the spider touches it with its forelimbs, equipped with chemoreceptors, as well as the so-called palpi (organs of touch), and, in addition, the spider tastes the victim with a bite.
For acquaintance with taxis in insects, we consider phototaxis as an example. These taxis (positive or negative) are found in insects primarily in the form of tropic, telo-, and menotaxis, although primitive taxis have not lost their meaning either. For example, positive and negative clinotaxis with clearly defined pendulum-like search movements play a leading role in the optical orientation of many fly larvae. The tropoxy and telotaxis are connected with each other by many transitional forms. In adult insects, however, distinct telotaxis are most common. As a rule, the tropotax movement soon turns into telotax movement. Mosquitoes and lice are dominated by the tropotaxis orientation movement.
Menotaxis belonging to more complex forms of orientation are found in higher insects (ants, bees, butterflies, beetles). The German researcher of the behavior of insects M. Lindauer highlights here the following four cases: 1) the angle of fixation of the light source is random (ants, beetle bug); 2) the angle of fixation is congenital (some butterflies); 3) the angle of fixation is reported by individuals of the same species to each other (“dances” of bees, informing about the orientation angle according to the position of the sun); 4) the angle of fixation is learned individually. The latter case can already be attributed to the highest category of taxis - mnemotaxis (learned orientation), which are widely distributed among vertebrates, but also occur in higher insects (bees, wasps, ants) (Fig. 37).

It is necessary, however, to emphasize that in those cases where orientation is based on other phototaxis, the behavior of insects is simultaneously determined by a number of additional factors. For example, it has been proven that in many insects the nature of their phototaxic behavior is strongly modified depending on air temperature, time of day, physiological state, etc. It is well known that a fly does not react to light while it flies quietly around the room. But it is worth starting to pursue her, as she is immediately “awakened” by a positive photothetaxis: she quickly goes to the light source, to the window.
In a similar way, insects react to a variety of mechanical, temperature, chemical and optical stimuli, to gravity, and, accordingly, they are found in many different forms of tigmo-, rheo-, anemo-, hydro-, geo-, thermo-, chemo-, photo - and other taxis.
Subject perception
Of particular interest is the ability of insects (and other arthropods) to optically perceive forms as a necessary component of the perceptual psyche. At the level of the elementary sensory psyche, the distinction of forms is still impossible.
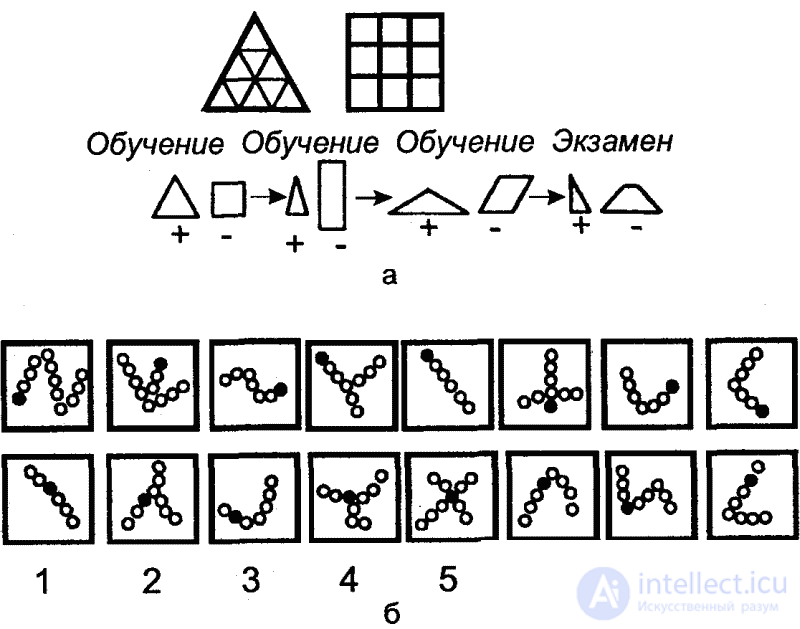
Until recently, it was believed that insects are capable of perceiving form, but only in specific frames. This specificity was seen in the fact that in experiments, insects, especially bees, were able to distinguish only figures resembling flowers in their outlines (for example, crosses or star-shaped figures), but in these experiments they did not distinguish simple geometric figures. However, Mazohin-Porshnyakov (in the works carried out jointly with G. M. Vishnevskaya) showed that bees can be fully successfully trained on such figures as a triangle, circle, square, from which he concluded that bees are able to recognize shapes directly by their graphic features.
Tinbergen's field experiments, in which female digging wasps were trained to recognize a circle of pine cones lined around the entrance to the mink, also testify to the ability of insects to recognize forms. After the wasp flew away for prey, the circle moved 30 centimeters to the side. After returning, the wasp looked for a mink first in the center of the circle (fig. 37). In the following experiments, the cones were also replaced with black stones, and a triangle or even an ellipse of cones lined up around the mink, but the wasp nevertheless flew in a circle, although it was known from previous experiments that the wasp was completely capable of distinguishing pebbles from cones. Thus, spatial orientation was carried out here only in form (circle).
Although in higher insects, the capacity for subject perception is beyond doubt, in this respect they are clearly inferior to vertebrates; Obviously, this is due to the specific structure of the organs of vision: the resolution of the eye depends on the number of ommatidia in insects. The more of them, the higher this ability, since the picture of the perceived object is divided into a greater number of points. In addition, insects, like other arthropods, obviously, are mainly oriented not by the substantive components of the environment as such, but by their individual characteristics, as is the case at the stage of the elementary sensory psyche.
In other invertebrates, which are also at the lowest level of the perceptual psyche stage, in cephalopod mollusks, vision plays no less, if not a greater role than insects, and is, of course, their leading reception (although these animals are oriented by well-developed touch and chemical sensitivity). Accordingly, the organ of vision received exceptional development in cephalopods: cephalopods have the largest eyes not only among the inhabitants of the seas, but also in general in the animal world, and not only in relative, but also in absolute terms. The eyes of giant octopuses (the genus Architeuthus) reach 40 centimeters in diameter with a total length of the animal (counting and “hands”) up to 20 meters. For comparison, you can specify that all, even the largest, whales (for example, a 30-meter blue whale) have an eye diameter of a little more than ten centimeters. The tremendous resolution (vigilance) of the cephalopod eyes is also striking: for one square millimeter, different representatives of cephalopods have from 40,000 to 162,000 rods, 120,000–400,000 for humans, and 680,000 for owls with the world's most keen eyes. As for subject perception, cephalopods, unlike lower mollusks, are already capable of genuine subject perception, which is expressed primarily in their discernment of the shape of objects. Thus, the octopus is able to distinguish a triangle from a square, a small square from a large one, a cross from a square, a vertical rectangle from a horizontal one, a triangle from a rhombus, etc. In total, it was established that an octopus can distinguish 46 different forms (B. B.'s experiments Boycott and J. Z. Young). Yet the cephalopod's vision, although approaching that of vertebrates, is also distinguished by a number of primitive signs that make the reflection of the objects of reality insufficiently full. There is still a lot of research to determine the specific features of the optical perception of the cephalopods.
Communication and group behavior
In higher invertebrates, the category of behavior is already fully represented, which, as has been shown, is only found in embryonic form in some representatives of lower multicellular invertebrates. We are talking about communication, which in higher invertebrates manifests itself sometimes in very complex forms of genuine group behavior (in contrast to colonial life, which is characteristic of many lower invertebrates). Recall that communication, in the exact meaning of this term, appears only when contacts between animals begin to occur through special signaling actions.
In cephalopods of mollusks and arthropods, especially in insects, we find clear communication systems with the transmission of information through different sensory channels. This applies primarily to bees and ants, which, of course, is associated with the complex, highly differentiated structure of their communities and the division of functions between the members of the latter (distribution of food-producing, protective and reproductive functions between individuals).
In ants, various forms of chemical sensitivity obviously play a larger role than other types of reception, and communication between individuals is carried out primarily through this channel: ant larvae emit specific substances that induce adult working individuals to feed them, by smell the inhabitants of one anthill recognize "their "And" strangers. " Even if an ant is alive or dead, its relatives will learn from specific secretions.
Coordination of the behavior of the inhabitants of the anthill occurs by sending and receiving chemical signals. It is enough to indicate the chemical means of ants (as well as bees and wasps), the selection of which serves as an alarm signal. These substances are released by ants in case of danger and quickly spread through the air. Their perception causes a defensive reaction in other ants (“soldiers”) (readiness to attack the enemy), although they are not yet aware of the source of the danger. Since, however, the range of such a signal, supplied by one ant (for example, in the event of an insect invading an anthill), is small, it can also indicate the place from which the danger is threatened. The greater this danger, the more ants emit chemical alarm signals and the more, therefore, individuals are called for help.
From the other excretions left on the substrate as a trace, the ants will know where, in which direction their congener has gone. In addition, the intensity of such an odorous trail indicates the amount of food found in the place where the trail leads, the more so that, going to the anthill, the ants put odorous marks along the way only when they found food. At the same time, the more tags are left, the more feed is found. As these tags evaporate rather quickly, the ants that follow this trail will perceive only fresh tags, and there will be fewer of these as the feed object approaches. Thus, the ants will eventually learn that the food resources in this place are exhausted, and will cease to go to it.
Some bees also have a chemical method of communicating information about food. This applies primarily to the bees - from the subfamily Meliponinae, who do not have the "language of dance." These bees, widespread in the tropics, have no sting, but they bite violently. In small forms (the smallest has a length of only 1.5 millimeters), the collector who found the food only brings other bees to an excited state, running restlessly along the honeycomb, pushing them and occasionally running up to the entrance. Departing bees are guided only by the smell of flowers. In other species, bee stimulation is initially achieved in a similar way, but additionally the picker, who brought bribes, causes the honeycombs to jolt into a vibrating state. After that, she returns to the detected flowers, and on the way back, leaves on the grass, branches or stones every two to three meters odorous marks, which then leads groups of other bees to the food source. This is achieved by the scout bee flying back and forth between two adjacent marks, until other bees join it. Consequently, here we are dealing with a complex transfer of information via optical, tactile-vibration and, possibly, acoustic channels, but the leading role is played by the odors left by the collector.
Our honeybee also has a chemical way of conveying information, but only as an auxiliary when it deals with food that lacks its own noticeable odor. In these cases, the location of the food object is marked with a secret from the so-called Nasonov gland.
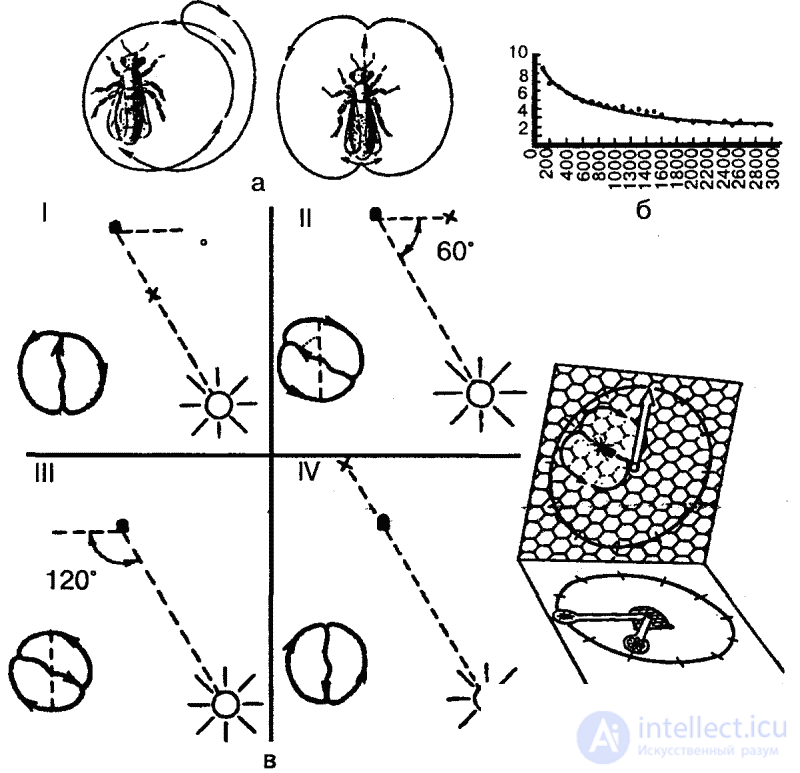
However, the main and most perfect way to transmit information about feed objects is “dancing”. Having found the source of food (nectar) and returning to the hive, the bee distributes to the other collector bees samples of nectar and proceeds to the “dance”, which consists of running along the honeycomb. The dance pattern depends on the location of the food source detected: if it is located next to the hive (2–5 meters away from it), then a “dance-push” is performed: the bee randomly runs along the honeycomb, wagging its belly from time to time; if the food is found at a distance of up to 100 meters from the hive, then one of the dances described by K. Frisch is performed - “circular”, which consists of jogging in a circle alternately clockwise and against it (Fig. 38). If nectar is found at a greater distance, then a wagging dance is performed, also described and studied by Frish. These are jogs in a straight line, accompanied by wagging movements of the abdomen, returning to the starting point either to the left or to the right (Fig. 38, a). Интенсивность виляющих движений указывает на расстояние находки: чем ближе находится кормовой объект, тем интенсивнее выполняется танец (рис. 38, б).
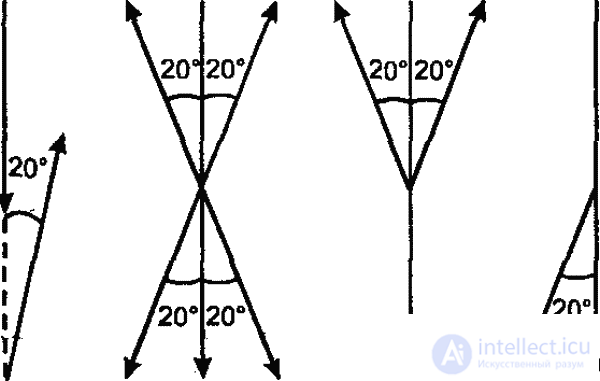
In addition, as Frish also showed, in the second dance form, the angle between the run line and the vertical (on vertical cells) corresponds to the angle between the bee's line of flight from the hive to the stern object and the position of the sun. Thus, the horizontal direction of the true flight is transferred to the vertical plane, the orientation of the position of the sun is replaced by the orientation of the earth's gravity. At the same time, if you should fly towards the sun, the bee dances from the bottom up, if from the sun, then from top to bottom (fig. 38, c). Thus, the honey bee is capable of transposing from photomentaxes to geomenotaxis: a fixed angle to the apparent position of the sun is transferred in a dark hive to gravity.
This ability of the honeybee to transpose, rooted in the widespread ability among insects to transfer menotax movements to the vertical plane (Fig. 39), ensures, as we see, the exceptional completeness and accuracy of information transfer. Such a transposition is a special case of transformation underlying all communication between animals: the properties of real objects and situations are transformed in the language of animals using a species-specific "code" into optical, acoustic and other conventional signals that inform other individuals about these properties. Despite all the complexity and accuracy, the communication system of the honeybee in principle does not differ from any other communication system in animals. Whether the true spatial parameters are translated into signals perceived by means of sight, touch and feelings of gravity, as is the case with a honeybee, or into signals perceived by chemoreception (ants), information is in any case transmitted in a transformed, conditional form. Here, the character and the functional significance of the psyche in animals appear in the best possible way.
The pinnacle of the development of instinctive components of communication is the ritualization of behavior, which is particularly clearly manifested in the reproductive behavior of insects, in particular in the "courtship" of males for females. Here is a typical mating "ritual" of a male marigold described by Tinbergen: after flying after the female, the male sits next to her, then stands in front of her, begins to "tremble" with wings (front-rear movements of closed wings), then re-opens and closes them, which accompanied by tapping the antennae on the wings of the female. After that, the male embraces the female with wings and pinches between them her antennae, which at the same time touch the odorous area on the wings of the male. Only after this, if the female does not reject the “grooming” of the male, mating begins.
In this example, attention is drawn to the strict fixity of the sequence of signal postures and body movements, which is typical of ritualized behavior. As we see, during ritualization, motor acts are always conditional: the movements performed by animals carry only a signal, but not directly “mechanical” function. It is on this basis that the language of the poses, body movements, and sounds in animals in all its amazing diversity originated. At the same time, the mental component of communication is directly embodied in the conditional signals.
The most complex systems of communication are inherent, of course, insects living in large families. Again, this applies primarily to the honeybee and ants. Differential and diverse relationships between members of the bee family, based on complex forms of communication, are unique in the animal world. A variety of opinions were expressed about this community, ranging from its anthropomorphic interpretation as a copy of human society or state (the unsuccessful, misleading name “social insects” was preserved) to the idea of a certain amount of independently acting “reflex automats”. At the same time, it turned out (the research of K. P. Trubetskoy) that a bee family is an association of individuals whose behavior is independent but not independent. The independence of individual bees is manifested primarily in pronounced individual differences: preference for certain types of hive work, differences in the timing and nature of the transition from hive activity to flight, etc. The relationship between the inhabitants of the hive is more complex in quantitative and qualitative aspects, resulting in a functional the mood of some bees can have an impact on others. Working individuals with their elevated levels of nervous activity often report arousal to other, less active worker bees.
Territorial behavior
Already in ringed worms and lower mollusks, the first manifestations of territorial behavior are observed, which manifests itself in the confinement of vital activity to a constantly occupied area, on which the refuge (“home”) of the animal is located. True, the performance of vital functions (or at least some of them) in a certain area constitutes only a part of territorial behavior, just as the presence of a “house” is not in itself a criterion of the latter. With full-fledged territorial behavior, which occurs only at the level of perceptual psyche, the host animal by its appearance, voice or smell makes its relatives know about their presence in a busy area. An important role is played by the “marking of the terrain”, that is, the application of odorous or other marks on different objects, especially on the periphery of the site.
Such marks can be feces or secretions of skin glands, as well as optical signs - bark ripped from tree trunks, trampled grass, etc.
A very significant sign of territorial behavior is the active protection of the landlord. Tinbergen attaches special importance to this criterion along with attachment to the locality. Hostility (“aggressiveness”) is manifested by the host towards any member of the same species, especially the same sex.
Often this behavior is confined to the breeding season. For example, a tree cockroach (Cryptocereus punctulatus), arranging a nesting chamber in rotten wood, very intensively (and usually successfully) protects the place where this chamber is located from the invasion of other males. In case of defeat, the nesting chamber, where the female is located, is taken by the winner.
In highly developed forms, territorial behavior, as shown by the German ethologist A. Heimer, is found in dragonflies. This is interesting only because dragonflies belong to the most ancient insects and, having appeared in the Paleozoic, have survived to this day, having not undergone significant morphological changes over these 50 million years. At the same time, as Haymer shows, the archaic structure is combined with highly developed behavior, and not only in the sphere of territoriality. This seemingly paradoxical fact of preserving the oldest signs of the structure in the presence of very progressive forms of behavior reaffirms the general rule of inconsistency between morphological and behavioral signs, bearing in mind the morphofunctional relationships at the level of the whole organism.
As for the territorial behavior of dragonflies (we are talking specifically about horned dragonflies-beauty Zygoptera), then mature males are looking for places suitable for permanent residence, which become their individual sections. These areas are labeled optically, namely by daily multiple fly-overs, and vigorously defend against male relatives. Inside the individual plot there are main and additional resting places, as well as a zone for laying eggs, the boundaries of which are marked in the mentioned manner. In this zone, the male leads the female by means of a “dance” performed together with her above the surface of the water. In addition, the male "cares" for the female, performing on its territory a special ritualized mating flight, and during egg laying, he guards and guards the female or several females, respectively, for one male often mates in a row with several females. The protection is manifested in the fact that the male drives away other males trying to mate with "his" females, which prevents the laying of eggs, while the guard function is that when the females attempt to interrupt this occupation, the male forces them to return and resume it. We also add that the territorial struggle of the males is carried out in highly initialized forms (there is no real struggle!). In general, all the behavior of males (as well as females) is distinguished by great plasticity.
Thus, in these species of dragonflies, there is a fully developed territorial behavior: individual areas are occupied in which functional areas are allocated (recreation, reproduction), in these areas all vital functions are fulfilled (except for sleep: in the evening males gather in special places of overnight stays), the host lets you know about your presence, marks and actively protects the site.
Interesting features of the territorial behavior of ants were identified by the Soviet entomologist A. A. Zakharov. It turned out that ants have two main types of use of feed areas: sharing of land by several families and use of the feed area by the population of only one nest. At the same time, a direct link between the ant density at the site and “aggressiveness” was revealed: in species with low density, the plots are not protected (except for the primordial zone), while at a high density, protected areas appear protected areas, and between them “neutral zones” (Fig. 40). As shown by another Soviet researcher, G. M. Dlussky, ants of the same species belonging to other families, as well as representatives of other species, are not allowed within these territories.
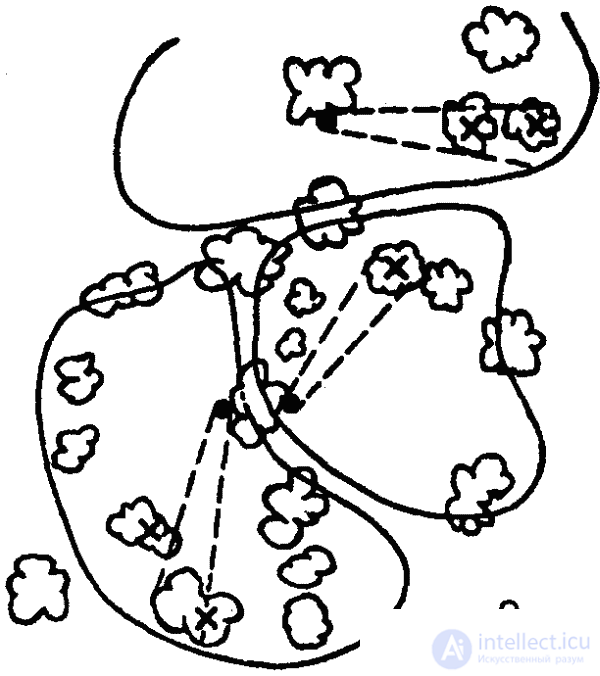
Territorial behavior has reached the greatest complexity in red forest ants, in which the highest density of individuals is observed in the feeding areas. The protected areas of these ants reach a much larger size than other species. The central elements of such a territory are ant trails (permanent feeding roads), the length of which often exceeds 100 meters, and the length of the entire network of trails is 1 kilometer. Each road is used only by a certain group of ants (a “column”), which occupies a certain sector in an anthill, adjacent to “its own” road. The territory of the whole family is divided by roads into separate parts that make up the territory of individual columns. Between them there are the same neutral spaces as between the territories of families, although not as pronounced as in the latter case. The boundaries of the territory are marked and prevent the “dispersion” of ants. Ants move through the territory not chaotically, but in accordance with the structure of the territory: in its central and middle parts - along the shortest path from the roads or the anthill, on the periphery - parallel to the borders of the territory. Thus, the trajectory of movement of each ant is determined by its location in the territory (research by I. V. Stebaev). The ants develop the greatest hunting activity on the periphery of the territory (further 10 meters from the nest). The collection of building material is made within the two-meter strip along the roads.
Zakharov rightly notes the similarity of the territorial behavior of ants with that of birds and predatory mammals.
Instinct and learning in the behavior of insects
For many years, the prevailing view was that insects and other arthropods were beings whose behavior was governed by a rigid "blind instinct." This view was rooted mainly under the influence of the works of the prominent French entomologist J. A. Fabre, who by his brilliant studies convincingly showed that even the most complicated actions of insects are not a manifestation of “reason”, but are carried out on an innate, instinctive basis. The one-sided development of Fabr's provisions led to the indicated, incorrect assessment of insect behavior, to the denial not only of the rationality of their behavior, but also to the denial, or at least diminishing of the role of accumulation of individual experience, learning in their life.
As we have already seen, the formation of any form of species-like, hereditarily “coded”, ie, instinctive, behavior in ontogenesis is always associated to some extent with any elements of individually acquired behavior, learning. Even relatively lower animals do not have to talk about strictly fixed instinctive behavior in a “pure form”.
This fully applies to insects, whose instinctive behavior is also improved by learning. This is the main role of learning in the life of insects. One can obviously assume that learning is in insects and other arthropods "in the service" of instinctive behavior. Like other animals, instinctive movements (innate motor coordination) are genetically strictly fixed. Instinctive actions as well, instinctive behavior are also plastic in insects to one degree or another due to the inclusion of acquired components in them.
Under natural conditions, the ability to accumulate individual experience is manifested in insects in varying degrees in different functional areas. Most often it is associated with orientation in space and food-processing activities. An example is the above-mentioned experiments on training bees for food reinforcement to orient themselves in different patterns. Another example is that ants, who very easily (in just 12–15 experiments) learn to go through even a complex maze, but, as far as is known, do not learn actions that lie outside the specified functional areas. Such a specific focus (and at the same time limited) ability to learn is a characteristic feature of learning from representatives of all types of arthropods.
The role of learning in the behavior of insects clearly appears in the "dances" of bees, these highest representatives of arthropods. Defending the view that insects, including bees, are "stimulus-related, reflex animals," American scientists V.Dether and E. Stellar say, for example, that bees do not learn to perform and interpret complex dance. At the same time, as shown by Soviet researchers N. G. Lopatina, I. A. Nikitina, E. G. Chesnokova, and others, learning processes not only clarify, but also modify the bee's communication abilities during ontogenesis and expand the set of signal means.
Moreover, as the aforementioned researchers have established, the biological significance of the signaling activity of honeybees is determined by the stereotype of conditioned reflexes acquired during ontogenesis as space is developed and during family communication. It turned out that the interpretation of the information transmitted in the dance about the distance and direction of flight to the food source is possible only if the bee has previously learned to relate the location of the feed to the nature of the information contained in the dance of the pickers. In addition, the tactile component of the dance (vibration of the abdomen) does not have an innate signal value. The latter is acquired in ontogenesis also by the conditioned-reflex way: the bees who had no contact (food) with the dancer during ontogenesis are unable to interpret this essential element of the dance. Therefore, each bee should basically learn to “understand” the language of dance. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность, — в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.
Of course, the honey bee occupies an exceptional position among insects, and far from all members of this huge class of mental development reaches such a height. Experimental data, testifying to the presence of analogues of some mental functions of higher vertebrates, testify to the exceptional mental qualities of a honeybee. We are talking about the highly developed ability of the bee to visualize generalizations, such as the “triangle” and “quadrilateral” types (regardless of the specific shape, aspect ratio, and relative orientation of the figures) (Fig. 41, a ), “bicolor”, etc. In one of the series of experiments, bees were asked to choose from pairwise presented figures those in which one local feature (a drawn circle) was at the end of a chain of circles, regardless of the length and shape of these chains (Fig. 41 b). With all the tasks offered to them, even in the most difficult versions, the bees coped well. At the same time, a large plasticity, non-standard behavior was noted, which the experimenter rightly associates with the continuous variability of environmental conditions (inconstancy of illumination, relative position, shape, color, and many other signs of environmental components) under which these insects have to obtain food. Mazokhin-Porshnyakov comes to the conclusion that the choice of an unfamiliar object on the basis of generalized visual images (sometimes incorrectly referred to as “concepts”) is evidence of the non-standard use of individual experience by bees, its use in a new situation, different from the situation of the initial development of the corresponding skill.
Thus, it is right to emphasize the presence and significance of the fact of transferring a certain skill to a new situation and solving a complex task based on individual experience, recorded as a generalized visual representation. In this regard, we actually find in psychical abilities psychic abilities similar to those related to the prerequisites for the intellectual actions of higher vertebrates. However, these prerequisites alone are not enough for intellectual behavior, animal thinking, especially if you look at these higher mental functions of animals as a step towards the birth of human consciousness. Therefore, the described abilities of bees cannot serve as a criterion for the recognition of their thinking, and in any case it is not necessary to speak of the rational activity of the bee, albeit in an elemental form, as Mazokhin-Porshnyakov interprets the results of his research. Recognizing the presence of peculiar thinking abilities and intellect in higher animals, one must be fully aware that reason, that is, reason, consciousness, as a qualitatively different category of mental reflection, is not inherent in any of the animals, but only to man.
General characteristics of the lowest level of perceptual psyche
At the lowest level of the perceptual psyche, all those progressive features that characterize the perceptual psyche in general are already represented, but in many respects the behavior of the animals belonging to it also carries primitive features that bring it closer to the behavior of lower animals. Thus, the main role is played by the orientation of behavior, as before, by individual properties of objects, but not by objects as such: subject perception clearly plays a subordinate role in general behavior. Equally, in the latter, rigid, “rigidly programmed” elements of behavior predominate over flexible, acquired, etc.
On the other hand, we state at this level a clearly pronounced active search for positive stimuli, that is, positive taxis behavior is developing strongly. There are all kinds of higher taxis, including mnemotaxis. The latter play an especially significant role in spatial orientation, and it is in the individual memorization of landmarks that the ability to acquire a change in species behavior and learning is manifested to the greatest degree.
At the same time, it is important to emphasize that although the animals considered here, in particular insects, accumulate individual experience, learning plays a significant role, there is also a certain inconsistency in the processes of learning, a combination of progressive and primitive features. The specific orientation, the confinement of these processes to certain functional areas, as well as the subordinate position itself, which is occupied by learning in relation to instinctive behavior, undoubtedly indicate the transitional position of a given level of mental development between the elementary sensory and developed perceptive psyche.
But, as we have seen, this in no way means that insects, like other members of the group of animals in question, lack plasticity of behavior. On the contrary, here too the general pattern is fully manifested, that the complication of instinctive behavior is inevitably combined with the complication of learning processes (and vice versa). Only this combination provides genuine progress in mental activity.
Instinctive behavior is represented at the level of mental development under consideration by already highly developed new categories: group behavior, communication, ritualization. Of particular difficulty are the forms of communication in species living in huge families, of which bees are best studied. The language of bees, these highest representatives of arthropods, is one of the most complex forms of communication that generally exist in the animal world. It is possible that new research will introduce us to astounding psychic abilities of other insects, but today bees seem to us the most developed in this regard. The most complex forms of instinctive behavior are naturally combined among them with the most diverse and complex manifestations of learning, which ensures not only exceptional consistency in the actions of all members of the bee colony, but also the maximum plasticity of individual behavior. The psychic abilities of bees (as well as some other higher insects) in some respects are obviously already beyond the limits of the lower level of perceptual psyche.
In a different direction than in arthropods, there was a development of mental activity in cephalopod mollusks. By some indications, they approached the branch leading to the vertebrates, as already indicated by their large size and the above-noted structural features of the nervous system and especially the visual receptor, which is directly related to a sharp increase in the speed of movement compared to other mollusks.
The behavior of cephalopods has not been completely studied, but many remarkable abilities are already known. First of all, they are distinguished by a significant complication of instinctive behavior. Cephalopods already have territorial behavior (occupation and protection of individual plots), “aggressiveness”, which is only planned for higher worms, group behavior (school life of squids and cuttlefish), ritualized forms of behavior appear in the breeding sphere, which is embodied in a species-specific “ grooming "males for females. All this is inherent only to higher animals, besides cephalopods, arthropods and vertebrates.
A number of researchers paid special attention to the “curiosity” that was very developed among octopuses, which is reflected in their examination of biologically “useless” objects, as well as their highly developed handling and constructive abilities. These abilities are manifested in the construction of shafts and shelters made of stones, shells of crabs, oyster shells, etc. This octopus picks up this building material, carries and strengthens it with its “hands”. Sometimes these nests are closed from all sides of the structure. According to some, though not yet confirmed, observations, octopuses are also capable of carrying weapons, using stones for protection.
Very important is the fact that for the first time cephalopods have the ability to establish contact with a person, to communicate with him, which results in the possibility of genuine taming of these animals (unlike insects!).
Thus, cephalopods, having left far behind other mollusks, as well as generally all lower invertebrates, have undoubtedly reached a high level of mental development, having in many ways become close to vertebrate animals.
However, in cephalopods there is the same inconsistency in learning ability as in insects. For example, an octopus generally has a well-developed ability to learn visual and tactile stimuli, but in some cases it is not able to solve seemingly simple tasks. This particularly applies to overcoming obstacles: the octopus is unable to find a workaround if the bait (crab) is located behind a transparent barrier (in a glass cylinder or behind a wire mesh); Vainly trying to seize the bait in the forward direction, the octopus lacks it from above, over the edge. True, according to Buytendijk, some octopuses are still capable of solving simple tasks of a workaround. At the same time, obviously, the former experience of the individual is of great importance. Other cephalopods are inferior to octopus in their mental abilities.
Of course, when evaluating such experiments, it is necessary to bear in mind that biologically inadequate and therefore unsolvable tasks are posed here: under natural conditions, an octopus never finds itself in a situation where the directly visible victim is inaccessible. In addition, the workaround tasks belong to the category of very complex - not only turtles, but also chickens cannot cope with them (in biologically incomparably more adequate conditions). Nevertheless, one should think that in the mental activity of cephalopods, progressive features that bring them together with vertebrates, and with primitive ones, a legacy of the lower mollusks, are indeed combined. The well-known "negativism" of learning also belongs to primitive features: cephalopods learn to avoid unpleasant irritations more easily than to find favorable ones. And in this it is not difficult to see a commonality with the behavior of animals that possess an elementary sensory psyche.
At the lowest level of perceptual psyche is, obviously, at least in part, a number of representatives of the lower vertebrates. However, the fundamentally different structures and lifestyles of arthropods and vertebrates are the reason that their behavior and psyche are essentially incomparable. So, one of the distinguishing features of insects are their small compared to vertebrate sizes. In this regard, the world around the insect is something quite special: it is not the microcosm of the simplest, but not the macrocosm of vertebrates. It is difficult for a person to imagine this insect world from his (from our point of view) microlandscapes, microclimates, etc. Although insects live side by side and with us, they live in completely different conditions of temperature, illumination, etc. For this reason, mental reflection In reality, insects cannot be fundamentally different than vertebrates, and most other invertebrates.
But since the most common signs of mental reflection, characteristic of a given level of it, are inherent in all the animals mentioned, it is possible, with regard to the insects we considered as an example, to say that we are dealing here with typical manifestations of a lower level of perceptual psyche, but in forms meet the special conditions of life of these animals, which were noted above.
The highest level of perceptual development of the psyche
In the animal world, evolution has led to three peaks: vertebrates, insects, and cephalopods. In accordance with the high level of structure and vital activity of these animals, we observe in them the most complex forms of behavior and mental reflection. Representatives of all three "peaks" are capable of objective perception, although, obviously, only in vertebrates, this ability has been fully developed. In the other two groups, perception developed in a peculiar way and qualitatively different from that in vertebrates. The situation with the other decisive criteria is similar with the stage of perceptual psyche, not to mention that only representatives of vertebrates reached the highest level of perceptual psyche in the evolutionary process, and that is clearly not all. Only in higher vertebrates are found all the most complex manifestations of mental activity that are generally found in the animal world.
Comparing invertebrates with vertebrates, it is also necessary to take into account that neither cephalopods nor arthropods have any relation to the ancestors of vertebrates. The path leading to these peaks deviated from the path to the third peak at a very early stage of the evolution of the animal world. Therefore, when compared with vertebrates, the high development of morphological and behavioral traits noted by us in these animals is only an analogy due to a powerful increase in the general level of vital activity characteristic of all three groups of animals.
From the phylogenetic point of view, echinoderms could be of much greater interest to us, which, like vertebrates, belong to the secondary rotates, unlike the primary rotates, in which the central nervous system is located on the abdominal side of the body and which, in particular, include mollusks and arthropods. However, modern echinoderms (sea urchins, stars, etc.) are not the ancestors of vertebrates, but only the lateral branch represented by highly specialized forms with primitive behavior characteristic of the elementary sensory psyche.
At this level, there are also lower chordates, which, together with vertebrates (or "cranial"), constitute the type of chordates. The lower chordates include the tunicates and the uncranial. Shells are marine animals, part of which leads a fixed life (asui-diya). Uncranial are represented by just two families with three genera of small marine animals, the most famous of which is the lancelet. In the uncranial and vertebral, a common feature is the internal axial skeleton, which has the form of a solid (lordlet chord) or articular (spine) rod, above which is located the central nervous system, which has the shape of a tube. Vertebrates are subdivided into classes of cyclostomes (lamprey and myxins), fish, amphibians, reptiles, birds, and mammals. Only the last two classes belong to higher vertebrates, within which, consequently, manifestations of higher mental abilities of animals are found. As for the lower vertebrates, there are outlined intermediate stages of mental development, characterized by different combinations of elements of the lower and higher levels of the perceptual psyche.
In vertebrates, the neural tube forms a swelling in the head section, turning into brain in the course of embryogenesis. Already the most primitive vertebrates, cyclostomes, have all five brain regions (medulla, posterior, middle, intermediate and forebrain). The process of differentiation and progressive development of brain structures, as is known, reaches its apex in mammals, and not only in the forebrain (the big hemispheres and their cortex), but also in the brainstem, where, in particular, centers of higher forms of instinctive behavior are formed. .
As for mammals, they generally occupy a special place in the evolution of the animal world, because, as Severtsov pointed out, “mammals very rarely adapt to rapidly advancing changes in the environment (for example, to new enemies, to new prey, etc.) by, that is, by slowly changing their organs and their functions. Much more often this happens by quickly changing old habits and skills and creating new ones adapted to new environmental conditions. Here, for the first time, a completely new and unusually important factor of the adaptive evolution of vertebrate animals, namely their psyche, appears on the scene. ” [53]
In the first and second parts of this book, the behavior of higher vertebrates was considered mainly. For this reason, we limit ourselves here to only a few additions to what has been said.
Locomotion
The vertebrate muscular system consists of segments symmetrically arranged along the body. True, the segmental arrangement of the muscles is less pronounced in chordates than in arthropods and worms, and in the higher representatives of vertebrates, it is strongly disturbed. There are somatic muscles that serve the effector organs, and visceral - muscles of the internal organs and skin. Somatic musculature always consists of striated muscle fibers.
The extremities of vertebrates are represented by unpaired and paired formations. Unpaired limbs are found only in cyclostomes and fish. These are the dorsal, anal and caudal fins. Парные конечности, передние и задние, сильно различаются у разных позвоночных соответственно своей функции (плавники, крылья, лапы, ласты, ноги, руки), однако сравнительное изучение строения их скелета позволяет отчетливо проследить эволюционные преобразования от общей примитивной начальной формы.
Основная функция конечностей состоит у всех животных в локомоции, в перемещении животного в пространстве. Однако у многих членистоногих и позвоночных сюда добавляется еще функция опоры приподнятого над субстратом тела. Поэтому в этих случаях говорят об опорно-локомоторной функции конечностей. Не вдаваясь здесь в детальное рассмотрение разнообразных форм этой основной функции конечностей позвоночных, укажем лишь на некоторые существенные моменты.
Видный советский ученый Н. А. Бернштейн писал, что постепенно назревшая в филогенезе потребность в быстрых и мощных движениях — привела на одной из его ступеней к возникновению и параллельному развитию «костно-суставных кинематических цепей скелета» и поперечнополосатой мускулатуры, снабженной соответственными нервными образованиями. У позвоночных эти скелетно-мышечные системы («неокинетические системы», по Бернштейну) получают существенное прогрессивное развитие по сравнению с членистоногими, причем особенно это относится к рассматриваемым здесь высшим позвоночным, в частности к их локомоторным способностям. Бернштейн указывает в этом отношении на глубокие качественные различия между низшими и высшими позвоночными, обусловленные усложнением возникающих перед организмом двигательных задач, возрастанием разнообразия реакций, требующихся от организма, более высокими требованиями в отношении дифференцированности и точности движений. «Достаточно напомнить, — пишет Бернштейн, — насколько, например, аэродинамический полет птицы сложнее почти полностью гидростатического плавания рыбы или насколько богаче по контингентам участвующих движений охота хищного млекопитающего по сравнению с охотой акулы. Молодая отрасль проворных теплокровных млекопитающих победила тугоподвижных юрских завров именно своей более совершенной моторикой». [54]
In modern ethological studies, locomotor activity is studied in its species-like manifestations as an adaptation to specific conditions of existence: the types and characteristics of walking, running, jumping, climbing, swimming, flying, etc., are determined by lifestyle features and are an important adaptation to the environment. At the same time, rhythmicity is inherent in all forms of locomotion, which is expressed in the fact that movements are performed in a clear sequence many times and in a relatively stereotypical way (although the behavior of the animal as a whole is not stereotypical). This rhythm is based on endogenous central nervous stimulation and proprioceptor feedback. In addition to proprioceptor sensitivity, external impulses only regulate these rhythms, correlate their parameters (force, speed, duration of movements, etc.) with the specific conditions of those situations in which the animal finds itself. In particular, external stimuli cause the onset or end of locomotor movements, although this may occur as a result of endogenous stimulation.
The foregoing sufficiently explains the fact that locomotor movements are among the most “automated” and uniformly performed components of the entire motor sphere of animals. In the same connection, the relative small number of forms of locomotion in each species stands. Determinant for locomotion is its physical, mechanical function. The locomotor movements themselves give the animal only minimal information about the world around it.
It is necessary, however, to take into account that locomotor activity includes indicative components, which, of course, have a certain cognitive value. So, for example, jumping animals, especially arboreal ones, must accurately "calculate" the distance before the jump. As shown by the Soviet researchers of animal behavior VM Smirin and O. Yu. Orlov, this is done with the help of special movements of “taking parallax” (fig. 42). Once in a new place, the flying squirrel “takes aim” at different objects, which she does before each jump, although with time the number of such movements decreases. As a result, the animal moving away from danger adheres to a previously “spent” path without unnecessary movements and makes jumps with amazing accuracy.

Manipulation
The effector organs of vertebrates always perform several functions at once — in addition to the main function, an even greater or smaller number of additional ones. This manifests the multifunctionality of these organs. As for the limbs of higher vertebrates, then, as already noted, their basis is the support-locomotor function: yet their additional functions are reduced to various forms of manipulating objects (food or inedible). Of particular interest to the zoopsychologist are forms of manipulation of the forelimbs, which in the process of evolution led to the primitive's instrumental activity and thus became the most important biological prerequisite for the emergence of labor practices among the most ancient people.
Speaking of manipulations, additional functions of the forelimbs in higher vertebrates, it is necessary to keep in mind that complex and diverse additional functions are inherent in these animals and the oral apparatus, and there is a far-reaching functional interaction between these two main effector systems. Therefore, it is advisable to analyze the additional functions of the front limbs and the oral apparatus in the complex. For example, in adult foxes, 45 additional functions of the forelimbs and mouth apparatus are found, in a badger - 50, in a raccoon - about 80, and in lower monkeys (macaques and baboons) - over 150 (Fabry data).
It is important to note that as mammals specialize in the field of locomotion, some of the additional functions of the forelimbs are transferred to the jaw apparatus. Thus, for example, in the series bear - raccoon - badger - fox, fox limbs are most suited for a long, fast run. But at the same time, the additional (manipulative) functions of the limbs are minimized and the corresponding vital movements (for example, in the field of nutrition) are performed almost exclusively by the oral apparatus. Just the opposite is the case with bears.
If we bear in mind the cognitive significance of manipulation, we should especially highlight those of its forms in which the animal holds the object in front of him freely on the weight with both or one limb. Such a fixation of the subject gives the animal the best opportunities for complex (optical, olfactory, tactile, kinesthetic, etc.) examinations, especially during the destructive impact on him (practical analysis). According to the data obtained by Fabry, this gives, under certain conditions, monkeys, and partly to other mammals capable of it, the most comprehensive and diverse information necessary for the development of higher forms of mental activity. It turned out that bears own three ways of fixing an object on weight, raccoons - six, lower monkeys and half monkeys - three dozen of such methods! In addition, only monkeys have sufficiently different motor capabilities to make a genuine destructive analysis (dismemberment) of an object on weight.
The formation of manipulative activity has already been mentioned (see part II). We only recall that the maxillary apparatus first begins to function, and later movements of the forelimbs appear, which, however, initially still need the support of a stronger system of the maxillary apparatus. Subsequently, the movements of the limbs become more and more powerful and independent and at the same time more and more diverse, that is, their multifunctionality increases. A number of functions, in particular, grasping, originally performed by the jaws, are transferred to the extremities. The peak of this development are the differentiated movements performed by only one limb. As already noted, the progressive development of additional motor functions of the front limbs was and is a major factor in mental development.
Comfortable behavior. Sleep and rest
A kind of manipulation can partly be considered a comfortable behavior that serves to care for the body of an animal, with the only peculiarity that the object of manipulation is not a foreign object, but its own body. But, in addition, this includes non-localized movements produced by the whole body and devoid of a special spatial direction. As a result, the following categories of comfortable movements can be distinguished: cleansing the body, shaking, scratching (a certain part of the body against the substrate), rolling on the substrate, bathing (in water, sand, etc.).
Comfortable behavior is widespread among arthropods (it suffices to recall how a fly cleans its paws on its head, wings, etc.), sometimes there are also special morphological formations or even organs for cleansing the body. The instinctive nature of comfortable movements is found in the species specificity of their performance, as well as in the strict fixation of body parts that are cleaned by one or another organ and method. Sleep and rest postures are just as clearly typical. So, for example, in bison, bison and their hybrids, a total of 107 types of typical postures and body movements were described in 8 areas of behavior (studies by M. A. Deryagina). The greatest number of poses (2/3) is in the areas of sleep, rest and comfort. At the same time, it is curious that in the pups (up to 2-3 months) of all three studied groups of animals, the species-typical differences in these areas are not yet expressed. The conclusion suggests that these differences are formed gradually at a later age.
If we consider one of the forms of comfortable behavior, licking, then, according to Deryagina, it turns out that only the bison licks the tail root or the raised front leg while standing, but the bison licks the back hips on the hind leg, and so on. Except for the hybrids, there are several There are only one ways of licking, of which only a bison is inherent, four are for a bison, and four are common for a bison and a bison. Here again there are clear differences in instinctive movements of closely related species (bison and bison are sometimes even considered as subspecies).
Sensory features
In full accordance with the highly developed effector systems of higher vertebrates, there is also a high level of their sensory abilities. The organs of hearing and balance are most important (the inner ear, starting with amphibians - additionally the middle ear, and in mammals also the outer ear), olfactory (olfactory bags and shells) and vision (eyeballs). Skin and muscular (tactile-kinesthetic) and thermal sensitivity, taste, and in some cases other types of sensitivity (electric, vibration, etc.) are also well developed. Correspondingly developed in vertebrates are various taxis.
Comparison of the role of sight and smell in the life of higher vertebrates shows that the majority of mammals play the leading role of smell, which reaches them with an amazing acuity (only sharks can be compared with them in this regard). Thus, the mole feels its prey through the thickness of the earth, the polar bear through the ice. The brown bear feels at a distance of 20 meters honey buried in the ground to a depth of 50 centimeters. The astounding olfactory abilities of dogs and ungulates are well known.
However, as already noted, in each particular case, the degree of development of the sense of smell (and the corresponding morphological structures in the receptor and brain) is determined by the characteristics of the biology of the species. For example, according to B. S. Matveev, even in related species living in similar conditions, but differing in biology, in the common vole (Mictorus arvalis) and the field mouse (Apodemus agrarius) there are significant sensory differences. Field mice eat a variety of food - a lot of seeds, as well as animal feed, while voles - mostly green food, which is always in abundance. As a result, the sense of smell is developed in a field mouse more than in a field mouse.
Vision is best developed in birds and primates. It plays an important role in their food-producing, defensive, reproductive and other forms of behavior, providing a clear orientation of the animal in space. Vision is of particular importance in the distant orientation of migratory birds, as well as in the intraterritorial orientation of mammals. For example, the orientation of birds according to topographic features (for example, the coastline), the polarized illumination of the sky, and astronomical landmarks — the sun and stars — have been well studied. The latter case is a clear example of orientation based on menotaxis.
In many mammals, orientation on their territory (individual plot or herd territory) is carried out using photo- and chemomnemotaxis (by optical and olfactory marks). In addition, even in worse-seeing mammals (for example, mustelids), vision plays an important role in targeting moving objects, especially when catching prey. Even animals with definitely poor eyesight, such as walruses, which are nearsighted over water, try to navigate with the help of not only smell, but also sight, especially again when dealing with moving objects (animals or people approaching them).
The most powerful sight, obviously, in the whole world of animals are birds of prey. Suffice it to say that a falcon is able, under favorable conditions, to see a sitting pigeon at a distance of almost one and a half kilometers. Equally, the vultures find their food (animal corpses) at a great distance by sight. But they are not able to detect the covered remains of animals. But this applies only to birds that inhabit the steppe expanses of the Eastern Hemisphere. Vultures of Central and South America (for example, black American vultures), searching for food in dense forests, cannot use sight and find it, however improbable it may seem, to be equally successful and from no less distance by smell. The situation is similar in mammals. In ungulates living in open spaces, vision is more developed than in their relatives living in the forests, even if the animals belong to the same genus (for example, bison is mostly a steppe animal and bison is an inhabitant of forests).
All these examples clearly show that the way of life, biology completely determine the nature of the behavior and psyche of animals (unlike a man who, thanks to his social and labor activity, achieved far-reaching independence from biological factors and whose mental activity is determined by the social conditions of his life). In the examples given, this provision of materialistic zoopsychology relates specifically to the sensory sphere of animals, in particular, to the question of the leading reception. As we have seen, in each individual case, even in closely related species, it is determined by the specific living conditions of the animal. But the same applies to mental reflection in general, the nature of which is also always determined by these conditions.
Visual generalizations and representations
The true reception, the true perception of the objective components of the environment as such is possible only on the basis of a sufficiently developed ability to analyze and generalize, for only this allows you to fully recognize the objective components of the environment that change their appearance (and other properties).
Studies conducted on different types of vertebrates (besides cyclostomes) have shown that they are all capable of objective perception, in particular, perception of forms. Already on this basis, we can conclude that all vertebrates are at the stage of perceptual psyche. However, within this stage, there are significant differences between the lower and higher vertebrates.
Thus, fish are able to experimentally distinguish geometric shapes, such as a square from a triangle, regardless of the changing size of these shapes. But in the control experiment it is enough to turn the triangle (point down) or square (to give it a diamond position), as the fish ceases to recognize these figures. Consequently, the perception and recognition of the form are limited here, not flexible enough. There is no ability to quickly transfer the formed visual representation on the basis of generalization. Mammals are quite capable of such a generalization, and they easily recognize a triangle or other geometric figure of any size in any position. Significantly superior fish and birds. True, the chicken, according to some sources, does not recognize an inverted triangle, which had previously been unmistakably distinguished from another geometric figure. However, the chicken is capable of significant generalizations, identifying, for example, geometric shapes, regardless of their color. Moreover, being trained to choose a solid triangle, it also reacts to it when it is presented to it in the form of a hatch or even only marked with three points! As well as fish, birds are able to recognize the relative size of the figures and also make mistakes on the basis of visual illusions.
The ability to visual generalize in many vertebrates has been experimentally proven. Especially with regard to their external representatives, it was shown that they are able, even in very difficult situations, to isolate essential details in perceived objects and recognize these objects in a strongly modified form. This suggests the conclusion that vertebrates have fairly complex general notions. Submissions are extremely important for survival. Individual experience is fixed in them in general, which allows an animal to more easily orient themselves in a significantly changing environment with a temporary absence of vital stimuli. Such visual generalizations are found, in particular, in the “extrapolation” phenomena described by L. V. Krushinsky, which are manifested in the fact that in some cases animals are able, following the movement of an object, to orient themselves on the expected path of its movement. Consequently, the forthcoming movement of the object after its disappearance is taken into account. In his experiments, birds and mammals were placed in front of the tunnel, which had a gap in the middle, through which it was possible to observe the movement of the feeder moving in the tunnel with the bait. In other experiments, a screen with a slit was used. Some of the experimental animals (corvids, predators) then ran around the tunnel or screen in the direction of movement of the bait and waited for it in the place of appearance. Such “anticipating” behavior, however, turned out to be capable only of animals that in natural conditions deal with hiding prey, which they have to hunt down or chase, often overtaking it, or themselves hiding food in reserve. As previously indicated, we are dealing here with the widespread ability among the higher vertebrates to find a workaround. It is important to note that in any case, when an animal goes around an opaque barrier or even simply turns away from the target object, it temporarily loses sight of the latter. Moreover, each representation by its very nature requires the absence of previously perceived objects.
Convincing evidence to the fact that generalized visual images in the form of representations are widely distributed among vertebrates has been given by experiments carried out by the method of delayed reactions (studies by V.S. Hunter, R. Yerks, O.L. Tinklpou, F. Boytendijk, V.Fishel, etc. .). In the Soviet Union, such experiments were carried out by N. Yu. Voytonis, I. S. Beritov, G. Z. Roginsky, and others. The usual scheme for conducting such experiments is this: the food hides in sight of the animal, which, however, allows him to find only some time later. Typically, the experimental animal must make a choice between several places where there may be a hidden bait. With a successful solution of the problem, there is a reaction to the missing stimulus, which is possible only if there are sensual ideas.
Experiments using the method of delayed reactions were successfully performed on various vertebrates, especially often on monkeys. The experiments that Tinklepaw conducted when the fruit (bananas) were hidden in front of the chimpanzees were very effective, and then, unnoticed by the animal, were replaced by a much less attractive food - salad or cabbage. Finding green after some time; the monkey was slow to take it and continued, sometimes with a shriek, a search. The latter lasted up to 33 seconds, while in the control experiment, when the monkey showed salad from the very beginning, which was then not replaced, the search continued for only 3 seconds and the monkey calmly ate it. The conclusion suggests itself that the monkey is looking for the object it initially saw, guided by a visual representation which, according to Tinklepow, also reflects certain quantitative and qualitative indicators.
Voytonis, on the basis of his experiments, also came to the conclusion that monkeys (macaques and chimpanzees) do not react to the feeder containing the bait, but to the food itself, more precisely, to a specific type of food. The same applies to dogs, but not to fish (Buytendijk and Fischel’s experiments).
The presence in vertebrates (and possibly some higher invertebrates) of ideas, expressed in delayed reactions and the ability to find workarounds (including extrapolation phenomena), gives their behavior exceptional flexibility and greatly increases the effectiveness of their actions at the search stages of behavioral acts. However, these abilities should not be overestimated, for they do not necessarily indicate a high level of mental development. Based on a developed memory, they are found in vertebrates at different phylogenetic steps depending on the animal's lifestyle: along with animals hiding reserves in different places, these abilities are characteristic of predators hunting hard-to-find, hiding, or just occasionally appearing animals.
This biological conditionality of the reaction in the absence of a stimulus that caused it before, was pointed out half a century ago by the Soviet zoologist and zoopsychologist D. N. Kashkarovna as an example of a cat that “stalks a mouse and sees that a mouse appears from a hole. The mouse disappears before the cat reacts. However, the cat gets up and goes to the hole. There can be no question that at the moment of reaction the determining stimulus is absent ... A cat could make the right choice between three holes only on the basis of the mouse’s immediately previous appearance, that is, on the basis of the idea of the latter. ” [55] As for monkeys, then, as can be seen from the mentioned studies, we are clearly dealing here with more complex complex processes.
The ability of higher vertebrates to subject perception, the ability to analyze and generalize, and thus to form ideas, is an important prerequisite for the development of complex skills that have already been considered before (see Part I) and which constitute the main content of the accumulation of individual experience not only in sensory, but also in the effector sphere, more precisely, in the motor-sensor sphere, acting as a single system.
Communication
In higher vertebrates, the processes of communication are of particular difficulty. Like other animals, the means of communication include elements of various modalities - olfactory, tactile.
Olfactory signaling, i.e., the transmission of information to other individuals by chemical means, prevails in territorial behavior, especially when marking the terrain of predatory and ungulates. For this purpose, special glands are used, located in different parts of the skin and emit a specific odorous secret. The latter is distributed through the air or applied to various objects (trunks and branches of plants, stones, etc.). Odorous tags are also produced with feces. The trace is marked with secretions of odorous glands, which contributes to finding each other by individuals of the same species. However, in all these cases, communication is not significantly different from what we observe, say, in insects.
Typical, instinctive components of the behavior of vertebrates, which are used for acoustic and optical communication between animals, are usually ritualized. Optical communication is carried out primarily with the help of expressive postures and gestures, as already mentioned above. Species-specific differences are often expressed in subtle to humans, but clearly genetically fixed details, causing very specific, often ritualized reactions in individuals of the same species. Especially often such differences are observed in closely related animals, primarily in signaling systems associated with reproduction. Here they serve as one of the factors of biological differentiation of species.
Specific forms of optical communication differ in higher vertebrates by a wide variety and differentiation. Often they are expressed in specific "dialogues" between two or more individuals, that is, in the mutual demonstration of body parts or poses. This particularly applies to “rituals” of intimidation or “imposition”. “Impression” expresses the superiority of one individual in relation to another, and has the tone of threat and challenge, but, in essence, is not aggressive in nature (as opposed to genuine “intimidation”, which is often followed by an attack). Often males "impose" on females during the "courtship" for them. The subordinate individual responds to the “imposition” of the “submissiveness” pose, which ultimately eliminates the possibility of collisions between animals.
In mammals, optical communication is often combined with the olfactory (Fig. 43), so that the allocation of communication systems for individual modalities is largely conditional in these animals.
This also applies to acoustic communication, as an example of which one should, of course, first of all recall the cries and songs of birds. Although the human ear is unable to capture all the sounds made by birds (much of them are in the ultrasound spectrum below 50 kHz), we are amazed at the huge variety of their voice responses. To the latter, it is necessary to add non-voice sounds, such as the whistling of wings during flight, tapping of a woodpecker on the trunk of trees, etc. The species specificity of such sounds makes them suitable for communication.

For a biologically adequate response to all these sound signals, the species-specific adjustment of the auditory system to a certain part of the spectrum is essential. As the Soviet researcher of bioacoustics of birds VD Il'ichev showed, the most important role in this setting is played by the peripheral parts of the auditory analyzer, which play the role of type-specific biological filters. With their help, there is also a special “adjustment” of the auditory system to the sounds that are especially important for an individual. Thus, the interdependence and unity of inborn functional (physiological and ethological) and morphological elements characteristic for the whole instinctive behavior clearly appear here.
The zoo-semantics of the sound signals of birds covers almost all spheres of their vital activity. These signals serve to identify an individual (in particular, its species), to alert other individuals about the physiological state of the expedition (this includes, for example, “begging”) or about changes in the environment (the appearance of the enemy or, conversely, the feed object, etc. .). At the same time, the rich zoopragmatic means of the bird allow the expeditioner to transmit to the perceptor highly detailed information. Thus, a rook flying up to the nest with feed, notifies the female incubating this with a shout that may have four different shades, to which the female also responds with different sounds. These shades of vocal reactions of birds are sometimes so thin that they are not perceived by the human ear.
Not only clear interspecific differences in acoustic communication, but also individual ones, by which individuals recognize each other (male – female, chicks — parents and each other, members of the pack — each other, etc.) are very important. It is interesting, for example, that, according to VG Thorp, in some cases, joint singing partners only respond to each other’s voices, but do not respond to the singing of other individuals. Especially individually different are the territorial cries of birds, announcing the employment area.
Большого разнообразия звуки достигают и у таких птиц, как куриные. Это обусловлено жизнью диких кур в труднопросматриваемых густых кустарниковых зарослях, где условия для оптического общения особенно неблагоприятны. Наиболее громкие и «впечатляющие» звуки куриных птиц — это крики петухов. Их слышимость (для человеческого уха) достигает 2 километров. В отличие, например, от звуков, с помощью которых петух подзывает кур к найденному им корму, кукареканье производится однократно, но зато дольше и на более высоких частотах. Это относится и к звуковым сигналам тревоги (например, при обнаружении врага), испуга или угрозы. Чем выше тональность, громкость и длительность этих сигналов, тем сильнее их эффект. Свое «кукареку» петух кричит на самых «высоких нотах», но всегда одинаковым образом: отклонения не превышают полтона.
Crowing is a kind of challenge to other roosters, and they respond with shouts of response, but only if rivals are roosters of the same “rank”. If a rooster occupying a subordinate position cries out within sight, then the dominant one simply attacks him, “not honoring” with a response cry. When the roosters, separated by a large distance and not seeing each other, have a common territorial screaming, i.e., acoustic marking of the terrain, warning about areas occupied by flocks of chickens - the rooster and its harem. The great biological significance of the cry lies in the fact that it prevents or at least reduces the frequency of cock fighting, and at the same time provides for the seizure, exploration and protection of the plots, and ultimately settling the species, but in a harmless way, because the case is limited to intimidation. The rival rooster is warned in advance that the site is already occupied and he will have to look for another suitable place for settlement. And only in the event that all such places are already occupied, will the battles for taking possession of the sites begin. In this case, the winner - the animal is stronger not only physically but also psychologically - will continue the race, and this, of course, is also beneficial for the prosperity of the species, for its progressive evolution. A similar role is played by other birds singing. In domestic chickens, the peasant's yard and the adjoining places remain for the rooster as his plot, the occupation of which he constantly recalls with a cry, although this behavior has already lost its biological significance, since man ensures all the needs of domestic animals and controls the evolution of the species. It is necessary to think that the extreme proximity of the location of the plots of domestic cocks makes them crow more intensely than it would have been in the wild, since in the village the population of chickens is “superdense”.
We do not dwell on the sound communication of mammals, since they are dominated by the same patterns as in birds.
Rigidity and plasticity in the behavior of higher vertebrates
As already mentioned, despite the still widespread opinion, instinctive behavior does not lose its significance in the process of evolution, since it cannot be replaced in principle by learning. We emphasize once again that instinctive behavior is a species behavior, learning is individual, and therefore the opposition of these two main categories of mental activity as supposedly different genetic steps is not justified. In fact, as we could see, the progress of instinctive behavior, in particular, in the series of vertebrates is inextricably linked with the progress of individually variable behavior, therefore vertebrates with highly developed forms of learning also possess no less developed complex forms of instinctive behavior.
At the same time, the rates of evolutionary transformations and the specific role in evolution of instinctive behavior and learning are different. Mainly, as Severtsov showed, this applies to higher vertebrates, especially mammals, in which the psyche acquires the value of a decisive factor of evolution due to the strong development of learning processes, including their highest manifestations - intellectual actions. But at the same time, the value of the instinctive basis of behavior is retained in full measure, as well as the slowly occurring changes in instinctive behavior. The given examples sufficiently show a great variety of true instinctive movements in higher vertebrates, which are quite typical, genetically fixed, and in some cases reach great complexity. Of course, in higher vertebrates, instinctive components serve primarily for the spatial-temporal orientation of the most vital behavioral acts. Spatial orientation is also carried out here on the basis of taxis: tropic, telo-, and menotaxis, that is, typical congenital elements of behavior, to which, however, are added mnemotaxes, characterized by the individual memorization of landmarks. In the latter case, we are already dealing with the accumulation of individual experience, and since it is the mnemotaxes that play a particularly important role in the life of higher vertebrates, then the innate methods of orientation are significantly enriched here by individually acquired methods.
In this regard, it should be noted that, like in other animals, the biological adequacy of the response to components of the environment is ensured in the higher vertebrates by the instinctive elements of behavior. Biologically significant objects are found in the surrounding animal environment in a very diverse and, most importantly, constantly changing species. We already know that their consumption or avoidance, that is, an adequate response to biological situations, is possible only if the animal is guided by some constant signs of these objects and situations. This is what happens on a genetically fixed, innate basis when an animal responds to key stimuli. This applies to higher vertebrates in the same way as to lower animals. But in the first reactions to key stimuli to a very large extent, they are concretized by individual experience; they are additionally oriented by well-developed subject perception. Thus, instinctive actions acquire a certain cognitive value for the animal, for they help it in acquaintance with the surrounding reality.
Instinctive behavior reaches a particularly high level of development in higher vertebrates in the ritualized communication of animals with each other. It is in the field of communication instinctive forms of behavior reach the greatest stereotype. It is absolutely clear that otherwise, without rigidly fixed zoopragmatic means, there can be no mutual understanding between animals, that is, there can be no genuine transmission of information. At the same time, it is full-fledged communication that is a prerequisite for higher integration in the field of behavior — integration of the behavior of individuals and entire communities.
However, even in the field of communication, the presence and importance of purchased components is beyond doubt. For example, in many birds, chicks will not be able to sing in a typical manner if they have not heard their parents' songs in time. The role of learning and in the formation of individual features of sound communication, as well as in numerous cases of acoustic imitation in adult birds, is undoubted. In the latter case, it turned out that the borrowed sounds can be used to communicate along with their own. On this basis, interspecific communication of birds develops, that is, the transfer of information between individuals of different species. Here, individually acquired communicative components with a clear signal value not only modify and enrich the actual instinctual components, but also act in a completely independent role.
Individually acquired elements of communication are of particular interest, since they show the possibility of going beyond the closed communication systems of animals. This especially applies to the communication of animals with humans, in particular, when training service dogs, etc. Probably, extended communication with animals played an important role in the domestication of wild species. All this, of course, was possible only at the highest level of the perceptual psyche, that is, in higher vertebrates. If we keep in mind the prehistory of anthropogenesis, which will be discussed below, then it must be admitted that the ability of higher vertebrates to expand their communication capabilities through learning should be an important prerequisite for the emergence of human forms of communication.
What has been said here about the role of acquired components in the instinctive behavior of higher vertebrates fully applies to other areas of behavior. Recall, for example, that bison and bison have the appearance of typical types of behavior only at a certain stage of ontogenesis. Although there is undoubtedly the maturation of certain types of typical motor components, regardless of the particular external conditions, the elements of learning are also involved in these processes. For the rest, we limit ourselves here to referring to what was previously said on this subject about the ontogeny of behavior.
There is also no need to say again that in different behavioral acts the proportion of innate and acquired components is different, as vertebrate skills contain qualitatively heterogeneous elements. We only recall that, not to mention the fact that skills are formed on the basis of unconditioned reflex processes, they always include conservative motor components that give skills in some respects similar to instinctive actions. Moreover, it is the formation of such conservative components that in many cases is the final result and the biological essence of the formation of a skill. It. refers to the motor stereotypes, automatisms, arising as a result of consolidation of the skill during training. Such primitive skills are found not only in lower vertebrates. On the contrary, learned automated actions play an important role in the life of higher mammals, including monkeys, as well as humans (for example, writing techniques, everyday "everyday" movements, etc.). In fish, on the other hand, such elementary skills are obviously limited to their ability to learn. This is connected with the fact that it is extremely difficult for fish, if it is at all possible, to alter a positive value of a stimulus into a negative one and vice versa. The reason for this is, in particular, the insufficient development of the associative functions of the brain of the lower vertebrates. As the Soviet physiologist B. F. Sergeev showed, only at the level of bony fishes and amphibians there appears the ability to form temporary connections between all analyzer systems and body functions, but there are still no intra-and inter-analyzer associative temporary connections. Analyzer systems are still fragmented, localized in different parts of the brain.
Only in birds and mammals, the switching function of the brain gets its full development, therefore complex skills, in which labile, flexible components are crucial, are found only in higher vertebrates. They determine the far-reaching variability, plasticity of the entire behavior of the animal, to which A. N. Severtsov attached such great adaptive significance. Indeed, complex plastic skills fully perform the function of the organism's rapid adaptation to transient environmental changes. The plasticity of higher order skills complements the rigidity of lower order skills and instinctive actions.
This plasticity is manifested in the possibility of rapid adjustment of the skill, in particular, when a positive or negative stimulus is converted into the opposite one. Another important feature is the ability to transfer skills to new conditions, in other words, the adequate use of accumulated sensory and motor experience with significant changes in environmental conditions. These possibilities are ensured by the strong development of plasticity in the sensory sphere, the ability for broad sensual generalizations, as already discussed above.
Let us explain this with a few examples. In the first experiment, the rat gets the bait as a result of a series of consecutive complex instrumental actions: it climbs the stairs to the platform, raises this ladder there with the help of a string thrown over the block, then climbs the ladder to the next platform and receives food reinforcement. Several more could be added to these phases - the essence of the matter does not change: we have here a complicated operation in which the sequence of individual actions of the animal is predetermined and strictly fixed by the conditions of the problem, i.e. those environmental components (obstacles) to which need directed activity of the animal.
In another example, with a relatively simple structure of an operation, the ability for far-reaching transfer of an operation under changed conditions is highlighted, which is also very characteristic of complex skills. In this case, the rat, having learned to find the shortest path to the “goal” in the maze, was able to do this without any new training even after the maze was set vertically. Since the animal is also guided by new sensations (from the vestibular apparatus), here the phenomena of transfer in the sensory sphere are evident.
In the third experiment, the rat, having mastered the land labyrinth, immediately sailed this labyrinth with no less success after it was filled with water. Since other movements are made during navigation than during walking, we can state here a far-reaching transference in the motor sphere while maintaining the same orientation in space.
Even in the first example, and even more so in the rest, it is impossible to regard the behavior of the experimental animal as a simple chain of mechanically learned movements, since in such a case the possibility of transferring the operation and preserving the skill under the changed conditions would be completely excluded. Difficult skills are exclusively dynamic motor-receptor systems that, based on highly developed orienting activities, produce highly flexible motor programs. The process of orientation merges here with the motor activity, and finding the right solution to the problem is formed in the course of this activity on the basis of highly developed sensory generalization.
The marked qualities of complex skills stand out among other types of learning, and it is thanks to them that complex skills have become the prerequisites and the basis for the development of higher forms of animal mental activity - intellectual actions.
Animal intelligence problem
Background and elements of intellectual behavior of animals
Intellectual behavior is the pinnacle of the mental development of animals. However, speaking of the intellect, the "mind" of animals, their thinking, it is necessary first of all to note that it is extremely difficult to indicate precisely about which animals one can speak about intellectual behavior, and about which animals - no. Obviously, we can talk only about higher vertebrates, but obviously not only primates, as it was accepted until recently. However, the intellectual behavior of animals is not something separate, out of the ordinary, but only one of the manifestations of a single mental activity with its innate and acquired aspects. Intellectual behavior is not only closely related to various forms of instinctive behavior and learning, but is itself composed (on an innate basis) of individually variable components of behavior. It is the highest result and manifestation of the individual accumulation of experience, a special category of learning with its inherent qualitative features. Therefore, intellectual behavior has the greatest adaptive effect, which A. N. Severtsov paid particular attention to, demonstrating the decisive importance of higher mental abilities for the survival of individuals and the continuation of the species in the presence of sharp, rapidly occurring changes in the habitat.
The prerequisite and the basis for the development of the intellect of animals - at least in the direction leading to human consciousness - is manipulation, especially with biologically "neutral" objects. Especially, as has already been shown, this applies to monkeys, for which manipulation serves as the source of the most complete information about the properties and structure of the objective components of the environment, because during the manipulation there occurs the most profound and comprehensive acquaintance with new objects or new properties of objects already familiar to the animal. In the course of manipulation, especially when performing complex manipulations, the animal experience is summarized, generalized knowledge about the objective components of the environment is formed, and it is this generalized motor-sensory experience that constitutes the main basis of monkey intelligence.
Pavlov said about the manipulation of apes with “biologically indifferent” objects: “This is the most persistent inquisitiveness. So the ridiculous statement that animals do not have it, is not in the germ of what we have and what ultimately created science, does not correspond to reality. ” [56] As an example, Pavlov referred to the manipulation of objects observed by a chimpanzee, in particular a box in which there are “no oranges, no apples”. Nevertheless, the monkey "has been busy for a long time ... on solving mechanical problems, which does not promise her any benefits, no material satisfaction." [57]
This, according to Pavlov, "pure, disinterested curiosity" and makes the monkey study the object of manipulation during the active influence on him. At the same time, different sensory and effector systems are included in the cognitive activity of the animal simultaneously and in interaction with each other. After all, the manipulating monkey almost continuously monitors the movements of its hands; under close visual control, a variety of actions are performed both without destroying the integrity of the object: turning in different directions, licking, stroking, crushing, rolling, etc., and of destructive order: breaking, tearing, isolating individual parts, etc. ( Fig. 24).
Destructive actions are of particular cognitive value, since they provide information about the internal structure of objects. When manipulating, the animal receives information simultaneously on a number of sensory channels, but in monkeys the combination of skin-muscular sensitivity of the hands with visual sensations is of paramount importance. In addition, smell, taste, tactile sensation of perioral vibrissae, sometimes hearing, etc. are also involved in the examination of the object of manipulation. These kinds of sensitivity are combined with the skin-muscular sensitivity of effectors (oral apparatus, forelimbs) other than monkeys and other mammals, when they manipulate objects. As a result, animals receive comprehensive information about the object as a single whole and possessing different quality properties. This is the meaning of manipulation as the basis of intellectual behavior.
It is necessary, however, to emphasize that visual perceptions, and especially visual generalizations, which have already been discussed, are of paramount importance for intellectual behavior. How developed is the ability to form generalized visual images even in rats shows the following experiment, in which rats successfully solved a very difficult task: an animal must choose from three presented figures (vertical and horizontal stripes) one dissimilar compared to the other two. The location and pattern of such a figure is constantly changing, therefore, it will be either vertical, then horizontal stripes, located either to the left, then to the right, then in the middle (in the wrong sequence). Thus, the experimental animal could be guided only by one, extremely generalized feature - the dissimilarity of one figure compared to the others. We have here, therefore, a case with visual generalization, close to the abstraction inherent in thought processes.
With the other element of intellectual behavior, this time in the motor sphere, we met when describing experiments with “problem boxes”. And here we are dealing with complex multi-phase skills, since in higher mammals, such as raccoons, it is relatively easy to achieve solutions to problems in which an animal has to open a set of various locking devices in a certain sequence. As with the described experience with tightening the rat ladder, a raccoon can solve this problem only if a strictly defined sequence of actions is observed. But the difference lies in the fact that, unlike a rat, a raccoon has to find this sequence itself, and this raises its activity, of course, to a higher level. True, according to some information, rats are capable of it.
It is necessary, however, to emphasize that even higher vertebrates solve instrumental problems more difficultly than locomotor ones. L. Kardosh noted in this connection that the cognition of spatial relationships prevails in the mental activity of animals (see Part I, Chapter 3), comprehended by them with the help of locomotor actions. In monkeys, especially apes, locomotor cognition of spatial relationships loses its dominant role due to the strong development of manipulative actions. However, only a person can completely get rid of the directing influence of spatial relationships, if this is required by the knowledge of temporal-causal connections.
The solution of multi-phase instrumental problems in monkeys has been studied by a number of researchers, in particular, N. N. Ladygina-Kots. In her monograph “The adaptive motor skills of the macaque under experimental conditions,” she summarized numerous experiments that used very diverse combinations of locking mechanisms. These experiments showed that the lower monkey (rhesus monkey) is able to learn to unlock large series of locking mechanisms, although it copes better with single installations. At the same time, there were numerous and diverse groping hand movements, “experimentation”. In view of her great haste, the adaptations that were not unlocked with light, quick movements turned out to be the most difficult for the monkey. Such movements as pulling, abduction, pulling, lowering, etc., were the easiest to do, the most difficult thing was moving away and turning, twisting. In general, in searching for points of delay and ways to overcome obstacles, the predominant role belonged to kinesthetic rather than visual perception. Interestingly, in many of these features, as shown by later studies, the actions of lower monkeys in experiments with locking mechanisms resemble those of raccoons.
An extremely important prerequisite for intellectual behavior is the ability to widely transfer skills to new situations. This ability is fully developed in higher vertebrates, although it manifests itself in different animals in varying degrees. V.P. Protopopov gives the following example of transferring acquired experience to a new situation in a dog. Initially, the experimental dog learned to open the latch on the door of the “problem cell” with the bait by pressing the paw. In other experiments, the same dog then learned to pull a piece of meat with its teeth and paws by the rope that lay on the floor in front of it. After this, a third situation was created that contained elements of the first two: on the cage used in the first situation, the latch was raised to such a height that the dog could not reach it with its paw, but a rope was tied to the latch, pulling which could be opened. When the dog was brought to the cage, immediately, without any other tests, she grabbed the rope with her teeth and, pulling, opened the latch. Thus, the task was immediately solved in the new situation, despite the fact that the previous elements were located in it in a completely different way: the rope was hanging, and did not lay horizontally on the floor, at the end it was tied not with meat, but with a heck, which was in another place - at the top. In addition, the heck was unlocked in the first experiments with the movement of a paw, and then with the help of teeth. "... A new skill," Protopopov writes about this, "is developed immediately," suddenly, "but this suddenness ... is caused by quite definite traces of past experience, which, under the influence of a stimulus, enter through a kind of explosive closure into a new temporary connection, and a new nervous structure and new reaction, different from the previous two, both in the receptor and effector parts ... Such skills ... may, by their external manifestation, imitate rational behavior and, if you do not know the stages of their occurrence, you can come to erroneous omorficheskim conclusions. " [58]
For example, N. R. F. Mayer, who recognized rats as capable of “reasoning”, arrived at similar anthropomorphic conclusions. The rationale for this conclusion was the results of his experiments on the development of delayed reactions in rats, during which these animals were able to bind elements of previous experience that had never been combined in their behavior before. As was shown, this happened in Protopopov’s experiments with the dog.
So, the ability of higher vertebrates to a variety of manipulations, to a wide sensory (visual) generalization, to solving complex problems and transferring complex skills to new situations, to full orientation and adequate response in a new environment based on previous experience are essential elements of animal intelligence. Yet by themselves, these qualities are still insufficient to serve as criteria for the intelligence and thinking of animals. Moreover, as mentioned, it is impossible to recognize such criteria as, for example, highly developed optical generalization abilities in bees.
The criterion of intellectual behavior of animals
A distinctive feature of the intelligence of animals is that, in addition to reflecting on certain things, there arises a reflection of their relationships and connections (situations). This partly has, of course, a place with some complex skills, which once again characterizes the latter as a transitional form to the intellectual behavior of animals. This reflection occurs in the process of activity, which, according to its structure, according to Leontiev, is two-phase.
We have already seen that the complex skills of animals are mostly multi-phase. However, these phases, be it the rat climbing up from the platform to the platform with the help of a pull-up ladder or the successive opening of the “problem box” closures, are essentially just a chain, a sum of unambiguous, equally-qualitative steps to solve the problem sequentially. As intellectual forms of behavior develop, the phases of solving the problem acquire a clear difference in quality: the activity, previously merged into a single process, differentiates into the preparation phase and the implementation phase. It is the preparation phase that is characteristic of intellectual behavior. As Leontiev points out, intellect arises for the first time where the process of preparing an opportunity to perform a particular operation or skill occurs.
In specific experimental studies, the biphasic intellectual actions are manifested, for example, in that the monkey pulls out the stick first, then with the help of this stick to knock a highly suspended fruit, as was the case in the widely known experiments of the German psychologist V. Köhler. In other experiments, the monkey could take possession of the bait only if at first it pushes it away from itself with a stick to a place where it (after a detour movement) can be reached by hand (Fig. 44).
Many other experiments were carried out in which monkeys had to solve a problem with the use of a tool (most often sticks). Thus, in the experiments of G. Z. Roginsky chimpanzees, who had experience with stick manipulations, they immediately used those to get the bait. But the lower monkeys, except for one (chakma baboon), were not immediately capable of this. Nevertheless, Roginsky rejects the opinion of V. Köhler on the existence of a gap between the psyche of apes and lower monkeys.
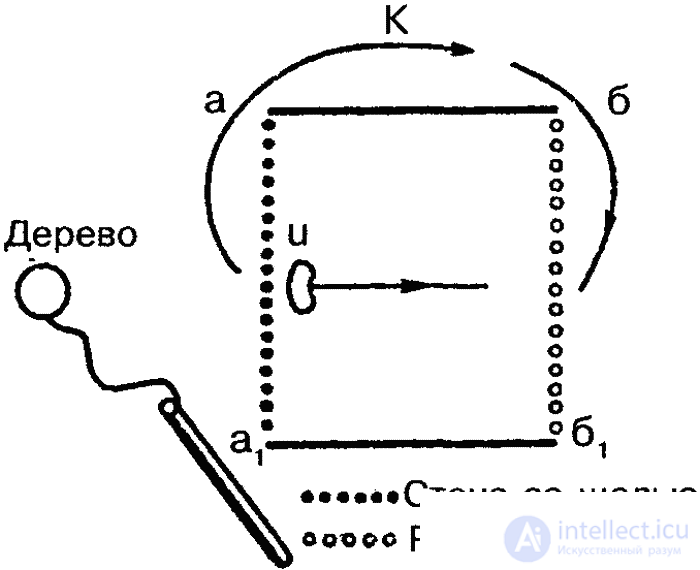
L. S. Novoselova, a Soviet zoopsychologist, was able with her research to identify the genesis of the use of sticks when solving complex problems in chimpanzees. She showed that the use of the stick is formed as an individually adaptive action, but is not a congenital form of behavior. At the same time, several stages are planned - from operating with the whole hand as a lever to specialized actions with a brush, which not only holds the stick, but also directs its movements in accordance with the specific properties of the instrument.
N. N. Ladigina-Kots studied in detail with chimpanzees the process of preparing and even manufacturing an instrument necessary to solve a technically simple task — pushing a lure out of a narrow tube. In the eyes of the chimpanzee, the bait was placed in the tube in such a way that it could not be reached with just fingers. Simultaneously with the tube, various objects were given to the animal, suitable for pushing out complementary foods after some “refinement” of them (Fig. 45). The experimental monkey completely (although not always immediately) coped with all these tasks.
In these experiments, the two-phase intellectual action also clearly emerges: the preparation of the instrument is the first preparatory phase, the pulling of the bait with the aid of the instrument is the second phase. The first phase, apart from the connection with the next phase, is devoid of any biological meaning. The second phase - the phase of implementation of the activity - is generally aimed at meeting a specific biological need of the animal (in the described experiments - food).
According to Leontyev, the first preparatory phase is prompted not by the object itself (for example, a stick), at which it is directed, but by the objective attitude of the stick to the bait. The reaction to this attitude is the preparation of the second phase, the phase of implementation, which is aimed at the object (“target”), which induces all the activity of the animal. The second phase includes, therefore, a certain operation, fixed in the form of a skill.
Of great importance as one of the criteria of intellectual behavior is the fact that when solving a problem, the animal uses not one stereotypically performed method, but tries different methods that are the result of previously accumulated experience. Consequently, instead of samples of different movements, as is the case with non-intellectual actions, with intellectual behavior there are samples of various operations, which allows to solve the same problem in various ways. The transfer and testing of various operations in solving a difficult task is expressed in monkeys, in particular, in that they almost never use tools in exactly the same way.
Thus, with intellectual behavior, we are dealing with the transfer of an operation, and this transfer does not require that the new task be directly similar to the previous one. The operation, as noted by Leontiev, ceases to be fixedly associated with the activity that meets a specific task. And here we can trace the continuity of complex skills.
Since the intellectual behavior of animals is characterized by a reflection not only of the objective components of the environment, but of the relations between them, the operation is carried out here not only on the principle of similarity of things (for example, obstacles) with which the operation was connected, but also on the principle of similarity of relations, connections of things with which she responds.
Forms of thinking
Based on many years of experimental research, Ladygina-Kots came to the conclusion that the thinking of animals always has a specific sensory-motor character, that this thinking is in action, and these actions are always subject-related. Such thinking in actions, according to Ladygina-Kots, is a practical analysis and synthesis, which, during the instrumental activity, is carried out in the course of direct handling of objects, in the course of their examination, processing and application. But at the same time they are not excluded, but, on the contrary, those generalized visual representations, which were discussed above, play a big role.
In accordance with this, Ladygina-Kots considers it possible to distinguish two forms of thinking that differ in complexity and depth (though only in apes).
The first form is characterized by the establishment of links of stimuli (objects or phenomena) directly perceived by the animal in the course of its activity. This is an analysis and synthesis in a clearly visible situation. An example is the choice of monkey items suitable for use as a tool, taking into account their size, density, shape, etc.
The second form is characterized by the establishment of links between directly perceived stimuli and representations (visual traces). Thus, in the experiments described, in which chimpanzees had to push a treat from the pipe with an instrument, mental operations of this type were manifested in the manufacture of an instrument, for example, in disentangling a ball of wire and straightening it. A particularly convincing proof of the existence of this higher form of thinking is the experience in which the monkey, along with the pipe, was given a plank much wider than the diameter of the pipe. The chimpanzee was able to completely separate the narrow torches from the board and use them as a tool to push the bait out of the pipe.
A monkey’s similar behavior can only be explained here by the fact that in the course of its previous activities a generalized visual representation of a stick-type object was formed (but only in the situation of the given task). The decisive role of previous experience in the formation of such visual "ideas" quite clearly appears in the same series of experiments. After all, even before the solution of the described task, the monkey was accumulating experience in the “refinement” of very different objects, including curly planks (fig. 46), which turned into a tool for chimpanzees no more difficult than breaking off side shoots (Fig. 45). However, the manufacture of tools from such a curly plank requires splitting off part of it in the longitudinal direction. Such actions and the experience accumulated at the same time prepared the monkey for solving the problem with a wide plank, which was given to it in one of the following experiments.
On the basis of establishing a connection between the generalized visual idea of the required object (such as a stick) and the direct perception of the second object (pipe), also given in a particular experience situation, the monkey managed to isolate (along an imaginary line!) A part of the whole — a splinter from the board, and thus that it turned out to be suitable to serve as a tool for pushing the bait.
Thus, chimpanzees are able to mentally decompose whole objects into parts, as well as complex figures into their constituent parts. As already mentioned, the leading role in the behavior, and in particular in the intellectual actions of monkeys, is played by their hands, the tactile-kinesthetic sensitivity of the hand. IP Pavlov therefore spoke with good reason about the "manual thinking" of monkeys. The combination of tactile-kinesthetic sensitivity with vision gives the monkey great advantages in establishing spatio-temporal relationships for practical analysis and synthesis. It was this significant expansion and deepening of the sensory sphere that formed in monkeys the basis of what IP Pavlov designated as “capturing the permanent connection between things” (or “the normal connection of things”). [59]
Biological limitations of animal intelligence
Along with all this, one must clearly imagine the biological limitations of the intellectual behavior of monkeys. Like all other forms of behavior, it is entirely determined by the way of life and purely biological laws, the framework of which even the smartest monkey cannot step over. So, for example, chimpanzees in the open each evening arrange themselves skillfully interwoven sleeping nests of branches and leaves, but, according to the testimony of the English monkey behavior researcher J. van Lavik-Goodall, they never build canopies and remain completely defenseless under the torrential tropical rain.
Very rarely do monkeys use in the wild and cannons. Правда, имеются отдельные наблюдения об орудийных действиях шимпанзе при добывании пищи или нападении. Но, как и другие человекообразные обезьяны, шимпанзе вполне обходятся в повседневной жизни без орудий. С другой стороны, другие позвоночные (каланы, галапагосские дятловые вьюрки и др.) систематически пользуются предметами в качестве орудий. Уже это указывает на то, что орудийные действия сами по себе не обязательно являются критериями высокоразвитой психической деятельности животных.
Биологическую ограниченность интеллекта антропоидов вскрывает и анализ экспериментальных данных. Так, Ладыгина-Котс показала, что зрительные образы, представления человекообразных обезьян являются значительно более слабыми, чем у человека, и всегда связаны с компонентами окружающей обстановки (ситуационная связанность представлений).
Эта ограниченность интеллектуального поведения неоднократно проявлялась в опытах Ладыгиной-Котс, когда шимпанзе допускал «нелепые» ошибки при употреблении предметов, предоставленных ему для выталкивания приманки из трубы. Так, например, он пытался втолкнуть в трубу кусок фанеры вопреки явному несоответствию ее ширины и принимался обгрызать его лишь после ряда таких неудачных попыток. Подчас неадекватность действий обусловливалась преобладанием деструктивного манипулирования (рис. 47). Кроме того, была проведена специальная серия опытов, в которых обезьяне давалась труба, закрытая с одного конца, и крючок. Если приманка, заложенная в такую трубу, была прикреплена к нити, шимпанзе легко вытягивал ее оттуда. Несмотря на это обезьяна не сумела адекватно применить крючок и, более того, чаще всего отламывала загнутую его часть как мешавший элемент. Ладыгина-Котс писала по этому поводу, что «шимпанзе оказался неспособным перейти от шаблонного привычного приема выталкивания приманки прямым, гладким единичным орудием к использованию приема притягивания к себе крюком», и усматривала в этом «недостаточную пластичность психики шимпанзе, ограниченность его мышления». [60]
Chimpanzees, according to Ladygina-Kots, “are not able to grasp at once the essential features in the new situation and establish new connections on the basis of comprehending directly perceived relations between objects”. [61]
This conclusion of Ladygina-Kots is confirmed by experiments and other researchers. So, the chimpanzee showed the situational connectedness of his ideas and the inability to embrace a significant change in the previous situation in the following experience: it is suggested that chimpanzees use an apple to stick the apple to the cage around the low wall. After the monkey has mastered this skill, part of the wall is removed immediately in front of the cage, as a result of which it would be more convenient to directly attract the apple with a stick. Nevertheless, the monkey continues to perform the same complex, difficult action, pushing the apple away from itself and circling it around the wall (EG Vatsuro’s experiment, Fig. 48).
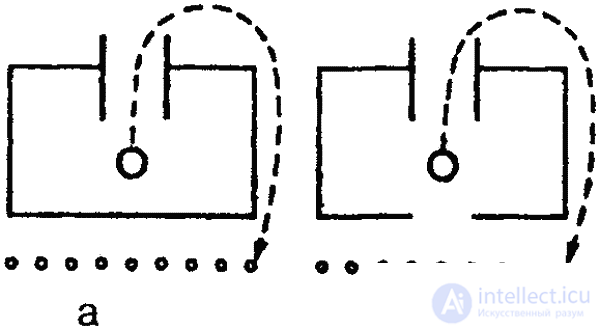
Even the most complex manifestations of monkey intelligence are in the end nothing but the use of a phylogenetically developed mode of action under new conditions. After all, the similarity of pulling a bait with a stick with pulling a fruit growing on a branch has long been noticed. Voytonis and Ladygina-Kots indicated that the developed ability of monkeys for practical analysis is connected with the peculiarities of their nutrition; highly developed sensorimotor functions of the hand, their combination with vision and, as a consequence, the distinctive cognitive abilities of monkeys, Fabry explains by the functional features of their grasping abilities (see Chapter 3), etc. This biological conditionality of all mental activity of monkeys, including anthropoids, is the cause noted the limitations of their intellectual abilities, the reason for their inability to establish a mental connection between representations alone and their combination into images. The inability to mentally operate with representations alone inevitably leads to an inability to understand the results of one’s actions, to understand true causal relationships. This is possible only with the help of concepts, which for monkeys, like all other animals, are completely absent for the above reasons.
In conclusion, we have to admit that the problem of the intelligence of animals has not been completely studied. Essentially, detailed experimental studies have been carried out only on monkeys, mostly higher ones, while other vertebrates have almost no evidence of experimental data on the possibility of intellectual actions. At the same time, as already mentioned, it is doubtful that intelligence was inherent only in primates.

Comments
To leave a comment
Comparative Psychology and Zoopsychology
Terms: Comparative Psychology and Zoopsychology