Lecture
A person perceives the world around him with the help of specialized sensory systems - analyzers. Perception is the process and the result of the formation of a subjective image of an object or phenomenon acting on the analyzer.
The classification of analyzers is based on the location of the receptors. Receptors are special sensitive nervous formations that perceive stimulations from the external or internal environment and process them into nervous signals. According to the localization of receptors, analyzers are divided into exteroceptive and interoceptive. In the first case, the sources of stimulation are outside the body; in the second, information comes from internal systems and organs. From the outside, the body is affected by visual, auditory, tactile, gustatory, olfactory stimuli, as well as the strength of the force (Table 5.1).
Table 5.1.
Exteroceptive analyzers and their characteristics
| Modality | Receptor localization | Type of receptors | Perceived quality |
|---|---|---|---|
| Vision | Retina | Sticks Cones |
Illumination Contrast Motion Dimensions Colour |
| Hearing | Snail | Hair cells | Height Sound strength Timbre Sound localization |
| Equilibrium | Vestibular organ | Macular cells | Rotation The force of gravity |
| Touch | Leather | Ruffini Finishes Merkel discs Taurus Pacini |
Heat Pressure Vibration |
| Taste | Tongue | Taste nipples at the tip of the tongue Taste nipples at the base of the tongue |
Sweet and sour taste Bitter and salty taste |
| Sense of smell | Olfactory epithelium in the nose | Olfactory receptors | Floral Fruit Musky Spicy scent |
Simultaneous and successive perception. All types of perception carry information about time, i.e. about the moment when the stimulus appeared and for how long it acted. In other words, perception is a process that starts with the moment when a stimulus acts on a receptor. The end of perception is the image of the stimulus - the object and its identification (identification). The duration of one act of perception can be very short, almost instantaneous, especially when the object of perception is well known. In this case, they talk about simultaneous (simultaneous) perception. If a person encounters an unknown stimulus, the duration of perception can increase significantly. It takes time to conduct a detailed sensory analysis, to put forward and test several hypotheses about the current stimulus, and only after that to decide what constitutes a perceived stimulus. In this case, they talk about sequential processing of information and successive perception.
Intensive study of the neurophysiological mechanisms of perception was made possible due to the emergence of methods for recording micro- and macropotentials of the brain, i.e. activity of individual neurons and the total bioelectrical activity of the brain. Perhaps, therefore, studies of the mechanisms of perception are numerous and include several levels of analysis: from a single neuron to a whole brain, and each level has its own version of the analysis of the perceptual process. However, regardless of the level at which perception processes are studied, one of the main places is occupied by the problem of coding. The main question is how reception and transformation of sensory stimuli takes place and how the perceived and transformed stimulus is reflected in the human CNS.
The teachings of Muller. The problem of transformation of information in the nervous system attracted the attention of researchers for a long time. The first ideas in this field were presented as early as the middle of the last century by Muller’s teaching about the specific energy of the sense organs. Its essence was that sensitivity to irritation does not depend on the stimulus acting on it, but on the properties of the excited nerves. For example, the optic nerve transmits the sensation of light, even if it is mechanically irritated (by a blow to the eye). Mueller and his followers believed that each sensation arises at the discharges of specific brain neurons that have their own "lines" of communication with peripheral organs. Various combinations of these elementary sensations should have created more complex types of perception. Of course, these ideas are mainly of historical interest.
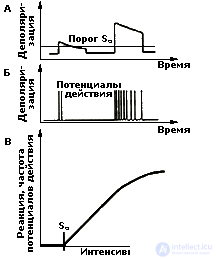
Principles of "Tagged line". At present, the physiology of sensory systems is very advanced (compared to other sections) the field of neurobiology, however the main problem, in fact, has not changed. How does impulse coming from specialized receptors of the senses transmit information of different types? The difficulty is compounded by the fact that, although the receptors are modally specialized and sensitive to a particular type of stimulation (sound, light, pressure, etc.), the nerves through which the impulses run are basically the same, and the impulses propagating from these receptors in the brain, have constant characteristics. (It is well known that a nerve impulse - action potential - is generated by a neuron on the principle of “all or nothing”).
The simplest answer assumes that the brain learns about the type of stimulus acting on the basis of the final destination in the cerebral cortex of the nerve impulses. Thus, action potentials entering the visual areas of the cortex carry information about visual stimuli, and similar impulses entering the auditory zones, about sounds, etc. In their fullest form, these ideas were embodied in the principle of the “tagged line”, according to which a direct morphological connection and, accordingly, the transmission of information from the receptor to a specific central neuron, which is responsible for determining the quality of the stimulus, is allowed.
However, how does the brain distinguish between the different qualities of each of the stimuli within the same modality, i.e. How does the brain differentiate between different visual or different sound stimuli? Such subtle distinctions are made on the basis of special forms of organization of the impulse activity of neurons, which are called codes.
Codes as a means of transmitting information. Information coding in the nervous system is the conversion of a specific energy of stimuli (light, sound, pressure, etc.) into universal codes of neutron activity, on the basis of which the brain performs the entire information processing process. Thus, codes are special forms of organization of the impulse activity of neurons that carry information about the qualitative and quantitative characteristics of the stimulus acting on the organism .
The problem of the formation of codes and their functioning in the central nervous system is currently the central core of the problem of presenting and transforming information in humans and animals.
From the point of view of one of the well-known experts in the field of sensory coding, J. Somien (1975) is most common in sensory systems to transmit information using the frequency of discharges of neurons. Other variants of neural codes are also possible: pulse density, intervals between pulses, features of the organization of pulses in a “pack” (group of pulses) - the periodicity of packs, duration, number of pulses in a pack, etc. There is a lot of data confirming that the listed characteristics of neural activity change in a regular way when the parameters of a stimulus change. However, the problem of coding is not limited only to the analysis of different variants of the pulse activity of neurons. It is much broader and requires more in-depth analysis (see Chrestomat. 5.1).
Currently, there are well-defined ideas about specific neural mechanisms that perform sensory analysis and the construction of a sensory model of the external environment. They are associated with the so-called concept of detector coding.
Detection concept. The main concept in the detector concept of coding is the concept of a neuron detector. A neuron detector is a highly specialized nerve cell that can selectively respond to a particular sign of the sensory signal. Such cells secrete individual signs in a complex stimulus. The separation of a complex sensory signal into signs for their separate analysis is a necessary stage in the operation of pattern recognition in sensory systems. Detector neurons were detected in the 60s. first in the retina of the frog, then in the cat's visual cortex, and subsequently in the human visual system.
Information about the individual parameters of the stimulus is encoded by the neuron detector in the form of the frequency of the action potentials, and the neuron detectors have a selective sensitivity with respect to individual sensory parameters.
Types of neuron detectors. The most detailed neuron detectors are investigated in the visual system. It is, first of all, orientation-oriented and direction-sensitive cells. For the discovery of the orientational selectivity phenomenon of the cat's visual cortex neurons, its authors D. Hubel and T. Wiesel in 1981 were awarded the Nobel Prize. The phenomenon of orientational selectivity lies in the fact that the cell produces a maximum discharge in frequency and number of pulses at a certain angle of rotation of the light or dark stripe or grating. At the same time, with other orientations of stimuli, the same cells respond poorly or do not respond at all. This feature gives grounds to speak about the sharpness of the setting of the neuron detector and the preferred response range. Direction-selective neurons respond to the movement of the stimulus, demonstrating a preference in choosing the direction and speed of movement (see Chrestomat. 5.2).
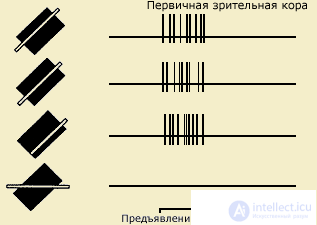
| A long and narrow strip of light causes the reaction of a complex cell, regardless of where it is presented in the receptive field, if its orientation is optimal (the three upper entries). If the orientation of the band differs from the optimal one, the cell reacts less or does not respond at all (lower record) (after Hubel, 1991) |
According to their ability to respond to the described characteristics of visual stimuli (orientation, speed and direction of movement), neurons-detectors are divided into three types: simple, complex and supercomplex. Neurons of different types are located in different layers of the cortex and differ in the degree of complexity and place in the chain of sequential signal processing.
In addition, detector neurons are described that react mainly to stimuli similar to those encountered in life, for example, moving hand shadow, cyclical movements resembling flapping wings, etc. This includes neurons that respond only to the approach and removal of objects. Neurons are also highlighted - color detectors. The most common neurons are sensitive to blue color (with a wavelength of 480 nm), green (with a wavelength of 500 nm) and red (with a wavelength of 620 nm).
In the higher centers of the brain, visual neurons are also found, which are particularly sensitive to stimuli similar to a human face or some parts of it. The responses of these neurons are recorded at any location, size, color of the "facial stimulus". It is important to note that these neurons are located not only in the neocortex, but also in the deeper structures of the brain - in the basal ganglia, thalamus, etc. In other words, among the external stimuli there are the most “preferable” ones, which are the most “convenient” for processing neural mechanisms of perception.
It is also assumed that there are neurons with increasing ability to synthesize individual features of objects and polymodal, i.e. with the ability to respond to stimuli of different sensory modalities (visual-auditory, visual-somato-sensory, etc.).
Neuron detectors are also described in other sensory systems: auditory and somatosensory. In the first case we are talking about detecting the position of the sound source in space and the direction of its movement. In the second, the activity of the neurons of the detectors is associated with the determination of the movement of the tactile stimulus along the skin or the size of the articular angle when the position of the limb changes.
Despite the fact that the available data on the mechanisms of detection in the visual, and especially in other modalities (auditory, somatosensory, olfactory) is clearly not enough, many researchers now consider the principle of neural detection as a universal principle of the structure and functioning of all sensory systems .
The generalized model of the sensory system. The detector coding principle underlies the "generalized model of the sensory system that performs active synthesis with the internal display of an external stimulus" (Sokolov, Vaitkyavichus, 1989). The model reproduces all stages of the information processing process from the onset of excitation at the receptor outputs to the formation of a complete image.
The transformation of the information flow in it is carried out using several types of formal neurons (detectors, gnostic neurons, modulator neurons, command, mnemic and semantic neurons), which are interconnected by two stable and plastic connections: informational and modulating.
It is assumed that the external stimulus through the senses creates a distributed excitation at the receptor output. As a result of the initial analysis, individual stimulus signs are distinguished from this excitation flow. At the next stage, the organization of the holistic image takes place, during this stage a hypothesis arises in the human visual system for individual fragments that this can be. Hypothetical representations of the object (the expected image) are extracted from the memory and compared with the information that comes from the sensory system. Next, a decision is made on the conformity or non-conformity of the hypothesis with the object, and the signs specifying the hypothesis are checked.
The concept of frequency filtering. As applied to the operation of the visual system, another supposed perception mechanism is described: frequency filtering. It is assumed that the visual system, primarily the cerebral cortex, adjusts itself to the perception of spatial information of a different frequency range. In other words, it is assumed that the human visual system contains neural complexes that are endowed with the properties of two-dimensional spatial-frequency filters that analyze the parameters of the stimulus according to the principle described by Fourier decomposition (see topic 2, item 2.1.1). Moreover, there appears to be a multitude of relatively “narrow” filters tuned to the perception of different spatial frequencies.
The advantage of a system based on frequency analysis is that it makes it easier to recognize familiar objects that are enlarged or reduced in size. It is assumed that only the harmonic composition is fixed in the memory system (the list of wave components obtained as a result of decomposition), it does not depend on the actual size of the object and this makes the identification of the stimulus more economical.
Debatable questions. Both concepts (detector and frequency filtering) do not give a clear answer to one question: how from a certain number of elements, whether they are reactions of neurons-detectors or Fourier coefficients, a complete image is created, i.e. the mechanism of visual synthesis and constancy of the visual image remains uncertain.
One of the founders of the detector concept D. Hubel characterizes this problem: "... often asked what are the most plausible assumptions about how visible objects are recognized, whether the cells become more specialized as they move to more central levels, so level can be cells that react to the face of a single specific person - for example, someone's grandmother? This view, called the "grandmother's cell theory," can hardly be taken seriously. M Do we find individual cells for a grandmother smiling, weeping or sewing? Or separate cells reflecting the concept or definition of "grandmother"? And if we really had "grandmother's cells", where would they send their output signals? Hubel, 1991. p. 228).
Neural ensemble as the basis of perception. As a possible solution, the following mechanism is proposed. The perceived object activates a certain group of cells - the "neural ensemble", each member of which may also belong to other ensembles. Since it is known that the destruction of a small area of the brain usually does not lead to the disappearance of certain memories, it must be assumed that the cells of one ensemble are not concentrated in one cortical zone, but are scattered in many areas.Thus, the “grandmother”, engaged in sewing ", must correspond to a larger ensemble, including the grandmother by definition, grandmother's face and the sewing process. The proposed scheme, however, is rather a hypothetical character.
Along with this, experimentally proved the existence of specialized neurons in the human brain. Numerous data of this kind were obtained in clinical studies of N.P. Bekhtereva with employees (1985, 1988). Moreover, they formulated a general psychophysiological principle, according to which the coding of the content of mental activity is carried out by combinations of the frequencies of impulse activity in the patterns of neuronal discharges of the brain and in the characteristics of their interaction. In particular, for example, it was shown that the patterns of the current frequency of neuronal discharges of some brain structures when perceiving verbal stimuli are able to reflect the acoustic and general semantic characteristics of words.
Nevertheless, in studies of perception performed by recording the activity of neurons, the problem of image formation and identification remains poorly understood (see Video).
The study of the physiological bases of perception is carried out using macropotentials of the brain: background electroencephalogram, as well as evoked and event-related potentials, and the use of EEG has two different aspects. The first is associated with the identification of EEG parameters related to stimulus coding. The second, more traditional, is the study of the EEG parameters of individual areas of the cortex or indicators of their joint interaction in the perception of stimuli.
ЭЭГ — показатели восприятия. Среди ритмов ЭЭГ наибольшее внимание исследователей в этом плане привлекает альфа-ритм (8-12 кол/с), который регистрируется преимущественно в задних отделах коры в состоянии спокойного бодрствования. Известно, что при предъявлении стимулов имеет место подавление или "блокада" альфа-ритма: причем она тянется тем больше, чем сложнее изображение.
Существует немало предположений, касающихся роли альфа-ритма в обеспечении активности мозга как целого. Основоположник кибернетики Н. Винер и вслед за ним ряд других исследователей считали, что этот ритм отражает механизм временного сканирования ("считывания") информации и тесно связан с механизмами восприятия и памяти. Действительно, по некоторым данным, биопотенциальное поле альфа-ритма связано с функцией сканирования информации в реальном времени и тем самым связано с процессами межмодальной сенсорной интеграции в процессах восприятия и памяти. По другим представлениям, альфа-ритм связан с действием селектирующих механизмов мозга, выполняющих функцию резонансного фильтра и, таким образом, регулирующих поток сенсорных импульсов.
Поиск электрофизиологических показателей восприятия в параметрах альфа-ритма опирается на представление о том, что воспринимаемая человеком информация кодируется комбинациями фаз и частот периодических нейронных процессов, которые находят свое отражение в характеристиках альфа-ритма. Предполагается, что такая особенность ЭЭГ, как пакет волн создается синхронизированной когерентной активностью группы нейронов, расположенных в разных участках мозга и образующих ансамбль. Предположительно все волны одного пакета хранят информацию об одном образе или его части и при восприятии опознается только тот образ, который закодирован ритмической активностью наибольшего числа нейронов в каждый данный момент времени (А.Н. Лебедев, 1985).
Экспериментально было показано, что значения параметров колебаний системы нейронов, оцениваемые, в частности, по расстоянию между соседними спектральными пиками внутри альфа-диапазона, могут служить аргументами уравнений, предсказывающих некоторые особенности восприятия. Например, чем больше период доминирующих колебаний в ЭЭГ человека и чем больше разнообразие воспринимаемых и ожидаемых стимулов, тем медленнее осуществляется их восприятие.
Наряду с этим существуют исследования пространственно-временных отношений потенциалов мозга при восприятии сенсорной информации. В отличие от предыдущего этот подход учитывает два фактора: время восприятия и его мозговую организацию. Речь идет о множественной регистрации ЭЭГ из разных зон коры больших полушарий в процессе восприятия. Поскольку нейронные ансамбли, участвующие в переработке информации распределены по разным отделам мозга, в первую очередь коры больших полушарий, логично считать, что перцептивный акт будет сопровождаться изменением пространственного соотношения ЭЭГ. Действительно изучение дистантной синхронизации биопотенциалов коры (М.Н. Ливанов, 1977) в ходе зрительного восприятия позволяет выявить следующее: вначале наблюдается преимущественная активация задних отделов коры обоих полушарий, затем в процесс вовлекаются передние отделы правого полушария. Узнавание связано с активным включением в процесс центральных и фронтальных зон коры.
Реорганизация биоэлектрической активности в процессе перцептивного акта характерна для всех видов чувствительности. Однако в зависимости от вида анализатора она имеет разную пространственную картину. Как правило, на ранних этапах процесса наибольшую активность демонстрирует проекционная зона (зрительная, слуховая, соматосенсорная), на завершающих этапах в процесс включаются передние отделы коры.
Вызванный потенциал как коррелят перцептивного акта. Наиболее адекватным инструментом изучения физиологической природы перцептивного процесса является метод регистрации вызванных (ВП), или событийно-связанных потенциалов. Последнее обусловлено тем, что, во-первых, ВП — это реакция мозга на внешнее раздражение, по длительности сопоставимая со временем перцептивного акта, во-вторых, ВП, взятые отдельно в каждой зоне коры, имеют свои характеристики, обусловленные функциональными особенностями этой зоны. Таким образом, при предъявлении стимула любой модальности (зрительного, звукового, тактильного и др.) в разных зонах коры возникают специализированные реакции, сопоставимые по длительности с реальным психологическим процессом восприятия.
Первые исследования в этом направлении ставили своей целью соотнести конфигурацию и параметры ВП с типом стимула и выделить варианты ответов, связанных с содержанием и физическими характеристиками стимула, например, установить отвечает ли мозг специфическим ответом на геометрические фигуры (круг, квадрат) или определенные слова. Предпринимались даже попытки определять по конфигурации ВП, какой стимул предъявлялся испытуемому. Несмотря на определенные положительные результаты, эти исследования не получили развития.
На первый план выступил иной подход. В его основе лежали представления о том, что компонентная структура ВП отражает не конкретное содержание или признак воспринимаемого стимула, а некоторые общие последовательные операции головного мозга по обработке стимула.
ВП и последовательная модель переработки информации. Было установлено, что в конфигурации ВП можно выделять компоненты двух типов: ранние специфические (экзогенные) и поздние неспецифические (эндогенные) компоненты. Обработка физических параметров стимула (сенсорный анализ) связывается с деятельностью специфических сенсорных систем и находит свое отражение преимущественно в параметрах экзогенных компонентов ВП. В зрительной модальности эти компоненты регистрируются в интервале до 100 мс от начала ответа, в слуховой и соматосенсорной модальностях — еще раньше, в пределах 10-50 мс от начала стимуляции. Эндогенные компоненты ВП, возникающие позднее и длящиеся до окончания ответа, предположительно отражают этапы более сложной обработки стимула: формирование образа, сличение его с эталонами памяти, принятие перцептивного решения.
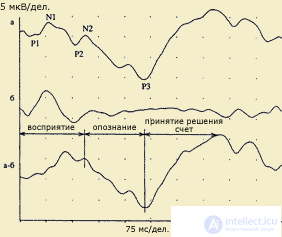
| Выделение когнитивной составляющей ответа: а - ответ в условиях опознания значимого стимула у здорового испытуемого (45 лет): P1-N1-P2 собственно ответ на стимул, N2 - 170 мс и P3 - 357 мс; b. Ответ на тот же стимул без опознания; а-б. Разность ответов в условиях распознавания и в стандартных условиях без распознавания. Когнитивный комплекс. Восприятие начинается на 76 мс, опознание и дифференцировка - компонент N2 - 170 мс, принятие решения и запоминание - компонент Р3 - 354 мс. (по В.В.Гнездицкому, 1997) |
Иллюстрацией такого подхода может служить концепция "информационного синтеза" А.М. Иваницкого (1986), согласно которой ранние компоненты ВП в интервале 0-100 мс связываются с оценкой физических параметров стимула, в среднелатентных компонентах в интервале 100-200 мс находят отражение процессы синтеза текущей сенсорной информации с содержанием памяти, а в поздних компонентах ВП отражается когнитивная оценка стимула как такового.
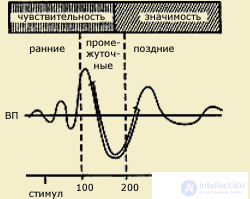
| Вызванные потенциалы (ВП) мозга, возникающие в ответ на световой или кожный раздражитель (по А.М.Иваницкому, 1999). Видно, что ранние волны соотносятся с чувствительностью, поздние - с мотивацией, значимостью стимула, а промежуточные - одновременно с двумя этими психофизическими показателями. Стрелкой на временной шкале обозначает момент предъявления стимула |
Эти представления хорошо согласуются с нейрокибернетическими построениями Д. Эделмена и В. Маунткасла (1981), согласно которым повторный возврат возбуждений является мозговой основой всех психических процессов. Так, например, момент возникновения образа совпадает по времени со встречей в коре двух информационных потоков, один из которых представляет собой внешний сигнал, а другой память. Предполагается, что эта память и есть возникающая в сознании человека частица "я", по отношению к которой сигнал воспринимается как нечто внешнее (см. Хрестомат. 5.3).
Изложенные выше представления объяснимы с помощью последовательной модели переработки информации, согласно которой каждая стадия перцептивного процесса наступает не ранее, чем реализуется предыдущая.
VP and other models of information processing. At the same time, there are data that are difficult to explain in the context of a sequential model of information processing. For example, when registering visual EPs for particularly complex stimuli, the parameters of these stimuli can be processed in the entire time range of the EP. The latter implies the existence of other models for processing information: parallel and cyclic. Moreover, within the framework of both models, VPs have different interpretations.
The parallel model is based on the idea of the existence in the visual system of several parallel operating subsystems in which the processing of information about different stimulus signs takes place independently, for example, the shape of an object is described using one channel, and the contrast, colors, etc. are used for another. In this logic, the components of the VI are considered as indicators of the inclusion of different channels.
Thus, the main difference between a parallel model and a sequential one is that the individual components are considered here not as markers for the implementation of specific stages of information processing, but as markers for the inclusion of different channels of information processing. For example, the component structure of EPs on chess fields and gratings is interpreted as a sequence of switching on different channels that process information about brightness (bright blue channel) and spatial contrast, the analysis of which is necessary to isolate the form (channel of form).
At the same time, differences in expressiveness relative to early and late IP components in responses to low and high spatial frequencies do not reflect the temporal sequence of processing of high and low spatial frequencies, but different spatial localization of EP generators to different spatial frequencies (Zislin, 1978). In principle, these assumptions have certain grounds, since they rely on electrophysiological data relating to sources of generation of EP, and they confirm that relatively early EP components on chess fields and lattices with a latent period of 75-125 ms are generated in the striatal cortex, and the later ones have source of the parastriar region.
The cyclical model suggests that in a perceptual process there may be separate and independent perceptual cycles with different internal structures and durations of individual stages. During cyclic information processing, apparently, the same neural populations are re-activated. Information processing at the same time is provided not only by single specific reactions of neurons, but also by their repeated involvement in the phasic process.
With a difficult perceptual task, the correct identification is more likely to occur at the later stages of information processing. As habituation increases, the probability of identifying a signal at earlier stages of processing increases. It is assumed that the number of CW components is determined by the number of information processing phases in cortical-subcortical neural networks (Trail et al., 1987). Thus, in the same time interval after the presentation of the same stimuli, there can be differences in the number of components and the configuration of the EP. Conversely, outwardly, the same VI components may correspond to different stages of stimulus processing.
These ideas cast doubt on the possibility of the existence of a fixed information-processing scheme of the type “constant stimulus - constant reaction”. At the same time, the logically consistent justification of the facts described above is obtained from the standpoint of the hypothesis of cognitive determinants of the organization of a perceptual act. According to this hypothesis, the sequence and set of operations (and the corresponding components of the EAP) in each perceptual act is not constant (from the sample to the sample) and is determined in the process of receipt of information and during its processing. The crucial role will be played by the conditions of the problem and the subject’s experience in the organization of information.

The basis of this approach is the idea of the systemic nature of the interaction of brain structures in the provision of mental functions (L. S. Vygotsky, A. R. Luria, E. D. Chomskaya, M. N. Livanov, O. S. Adrianov, etc.) . The problem of the involvement of different parts of the brain, primarily the cortex, in providing perception is studied experimentally using various methods: electroencephalography and evoked potentials, computed tomography, direct stimulation of the cerebral cortex, analysis of perception disorders in focal brain lesions.
The functional provision of perception is connected with the activity of the second block. Morphologically, it is represented by the posterior divisions of the cerebral cortex and includes “apparatuses” of the visual (occipital), auditory (temporal), general sensitive (parietal), and also somatosensory (postcentral) zones of the cortex. All the listed “devices” are built according to a general principle: they include primary projection zones (“cortical ends of the analyzers”) and associative secondary and tertiary zones.
A common feature of primary zones is topical organization, i.e. each area of such a zone corresponds to a certain area of the peripheral receptor surface. The projection of the retina forms the retinotopic organization of the occipital zone, the projection of the organ of Corti is the tonotopic auditory zone, the projection of the receptor surface of the skin is the somatotopic somatosensory zone. This principle of organization provides a kind of mapping of the retina, Corti organ and skin surface.
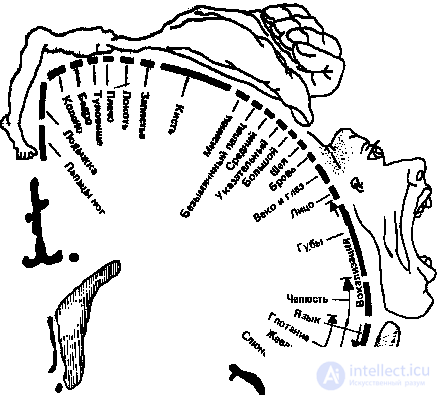
The somatotopic organization of the somatosensory zone has been studied best of all. A well-known somatosensory homunculus is a schematic "little man" reflecting the spatial representation of different parts of the body in this zone of the cortex. In fact, it presents a map of this zone, which was obtained during operations on the human brain using direct electrical stimulation. Such stimulation causes sensations in a person, which are distributed over different parts of the body, but unevenly — the functionally most significant areas of the receptor surface (face, tongue, hand) are most represented in the cortex.
The latter is true for all projection zones, for example, in the occipital zone, most of it is occupied by the projection of the central section, where cones are predominantly concentrated, providing high contrast sensitivity and color vision.
Thus, when the stimulus is perceived, the entire projection zone is not evenly excited: mainly those neurons that receive information from excited peripheral elements, the receptors, are activated. This primary stage is often referred to as sensory analysis.
The nature of information processing changes fundamentally during the transition to the secondary associative zones of the cortex. Being modally specific, these zones, according to the figurative statement of A.R. Luria, transform topical projection of irritation into a functional organization. This means that information processing in associative zones occurs on a different basis. Based on the ideas formulated in Section 5.1, it can be assumed that when information passes from the primary projection zone to the associative, the nature of the codes in which the information is processed changes. Due to which mechanisms exactly the reorganization of the neuron impulse activity flows is carried out is still unknown. Above (see p. 5.2) it was said about the existence of neurons of different specializations (mnemonic, semantic, gnostic, etc.), neural ensembles performing generalization functions in perception. However, there is not enough experimental data to describe the neural processes that accompany the processing of information in the secondary areas of the cortex. Nevertheless, the activity of these zones is associated with the integration of disconnected excitations of the primary projection cortex and the formation of complete images of perception. The functions of the tertiary zones are associated with the processes of intermodal generalization and construction of the picture of the world.
Morphofunctional levels and stages of information processing . The processing of information has many stages and begins at the receptor level: the sensitive elements of the skin, retina, Corti organ in the cochlea, etc. In addition, each analyzer includes a number of subcortical switchings - links (in particular, for example, at the level of the thalamus), where the initial processing of information is carried out. Only after these stages (that is, carried out at the subcortical level), already partially processed information enters along the pathways into the projection zones of the cortex and from them into the secondary and tertiary ones. This overall sequence of stages has its own specifics for each type of sensitivity.
So, in the visual analyzer there are two systems for processing information about the outside world. The first of them is responsible for identifying the object, the second determines the localization of the object in the external visual field. Performing different functions of visual perception, these systems differ in their morphological structure. Although both systems start in the retina, the first system originates from the so-called type X cells, the second system starts from the U type cells. Further, these systems have their representation in different subcortical centers: the first system continues processing information about the shape of the object in lateral articulated bodies the thalamus, from where information enters the visual centers of the cortex: the primary projection zone, the secondary associative fields, and from there to the lower temporal zone of the cortex. When selectively affecting certain areas of the above zones, the perception of individual qualities of an object suffers: only colors, only forms or movements. Integration of all separately processed features of an object occurs in the lower temporal cortex, which is responsible for the final formation of the integral visual image of the object.
The second system has a different brain topography. From the retina, the fibers of this system are sent to their own subcortical centers - the upper double-flanks. In this structure, it is not only the visual perception of the spatial characteristics of the object, it also contains the centers that control the movement of the eyes. The activation of these centers triggers the saccade — a rapid, abrupt movement of the eyes, the amplitude and direction of which ensures that the stimulus hits the central visual field. This is necessary because it is the central visual field that possesses the best abilities to perceive high-contrast complex stimuli. Further processing of information in this system is associated with the activity of another thalamic nucleus, the so-called pillow and parietal region of the cerebral cortex. In this area, information is integrated from the primary visual cortex and centers that control eye movements. The merging of two information flows creates a constant, i.e. permanent screen of the external visual field. Due to this, the image of the visual world moving along the retina while the eyes are moving remains unchanged.
Thus, on the basis of the interaction of information coming from different parts of the visual system and the system of control of the oculomotor activity, a holistic and stable picture of the worlds is being built.
A model of visual perception, taking into account the morphological structure of the analyzer, suggested V.D. Glezer (1985). He distinguishes four levels of processing of visual information: from the visual centers of the subcortex, implementing elementary sensory analysis, through the projection cortex, where spatial-frequency analysis is carried out, to the third level, at which the segmentation of the visual space into individual figure elements occurs, and from it to to the higher visual centers of the lower temporal and late-dark areas of the cortex, which carry out identification and training, this is the way of transforming and describing visual information according to the Glezer scheme.
The proposed model is not the only one in the description of the proposed mechanisms that ensure the integrity of visual perception. The problem is that each analyzer has vertically organized links (located on the path from the receptor to the primary projection zone), horizontally organized links (primary, secondary and tertiary zones of the cortex) and laterally organized links (symmetric subcortical formations and zones of the cortex in the left and right hemisphere). However, in most cases, the proposed models are limited to one or two of the listed components.
Perceptual specialization of the hemispheres. The features of the functional specialization of the left and right hemispheres of the brain are widely studied. A detailed analysis of this problem is presented in the books of N.N. Bragina and T.A. Dobrokhotova (1981, 1994), E.D. Chomskoy (1987), S. Springer and G. Deutsch (1983) and many others.
The human nervous system is designed in such a way that each hemisphere of the brain receives information mainly from the opposite side of the body. This principle of contralateral projection refers both to the general bodily, tactile sensitivity, and to sight and hearing, although with respect to the latter the picture is not so unambiguous. However, the use of appropriate methods allows to selectively provide information only in one hemisphere and thereby identify significant differences in the functional abilities of the two hemispheres.
The most studied hemispheric relationships with visual and auditory perception. Features of the specialization of the hemispheres in the perception of sound stimuli, first of all speech, are presented in topic 8 (see § 8.4; § 8.5), therefore, the main focus here will be on the specialization of the brain hemispheres in visual perception.
As shown by clinical studies, the destruction of the central visual zones in one of the hemispheres leads to the loss of the opposite half of the visual field (right for left-sided damage and left for right-handed). Knowing the site of damage to the visual cortex, one can predict what the vision loss will be. However, it is difficult to predict the human response to such damage, because the human nervous system has compensatory capabilities.
Among the hypotheses regarding the nature of inter-hemispheric differences in visual perception, two are common: one connects these differences with the verbalization of perceived stimuli, on the other, the differences are rooted in the peculiarities of the working style of each hemisphere - analytical for the left and holistic global for the right. In general, the point of view dominates that the superiority of one or another hemisphere in the perception of visual stimuli is determined by the ratio of two stages of processing: the visual-spatial, in which the right hemisphere prevails, and the verbalization processes implemented by the left.
Attempts are being made to explain inter-hemispheric differences based on the specialization of neurons in the perceptual process. Known in this regard is the model that ascribes a specialization to the cerebral hemispheres regarding the processing of different spatial frequencies. According to these assumptions, frequency filtering is carried out in different ways in the visual centers of both hemispheres. The right hemisphere with great success handles low spatial frequencies and, therefore, is more associated with the perception of the contours of objects and their large parts. The left one, on the contrary, handles high frequencies more successfully, and is more responsible for the perception of fine details of the image.
In the most general form, the specialization of the hemispheres in providing visual perception is given in table 5.2.
Table 5.2.
The differences between the hemispheres in visual perception
| Left hemisphere | Right hemisphere |
|---|---|
| Incentives are better recognized | |
| Verbal Easily distinguishable Acquaintances |
Nonverbal Difficult to distinguish Unfamiliar |
| Better perceived tasks | |
| Evaluation of temporal relationships Establishing similarities Identification of incentives by name Transition to Verbal Coding |
Evaluation of spatial relationships Establishing differences Establishing the physical identity of the stimuli Visual spatial analysis |
| Особенности процессов восприятия | |
| Аналитичность Последовательность (сукцессивность) Абстрактность, обобщенность, инвариантное узнавание |
Целостность (гештальт) Одновременность (симультанность) Конкретное узнавание |
| Предполагаемые морфофизиологические различия | |
| Фокусированное представительство элементарных функций | Диффузное представительство элементарных функций |
В целом следует заключить, что правое "пространственное" и левое "временное" полушария обладают своими специфическими способностями, позволяющими им вносить важный вклад в большинство видов когнитивной деятельности. По-видимому, у левого больше возможностей во временной и слуховой сферах, а у правого в пространственной и зрительной. Эти особенности, вероятно, помогают левому полушарию лучше отмечать и обособлять детали, которые могут быть четко охарактеризованы и расположены во временной последовательности. В свою очередь единовременность восприятия пространственных форм и признаков правым полушарием, возможно, способствует поиску интегративных отношений и схватыванию общих конфигураций. Если такая интерпретация верна, то, по-видимому, каждое полушарие перерабатывает одни и те же сигналы по-своему и преобразует сенсорные стимулы в соответствии со специфичной для себя стратегией их представления.

Comments
To leave a comment
Psychophysiology
Terms: Psychophysiology